

Molecular basis for the RIN4 negative regulation of RPS2 disease resistance
- PMID: 15749765
- PMCID: PMC1088003
- DOI: 10.1105/tpc.104.030163
Molecular basis for the RIN4 negative regulation of RPS2 disease resistance
Abstract
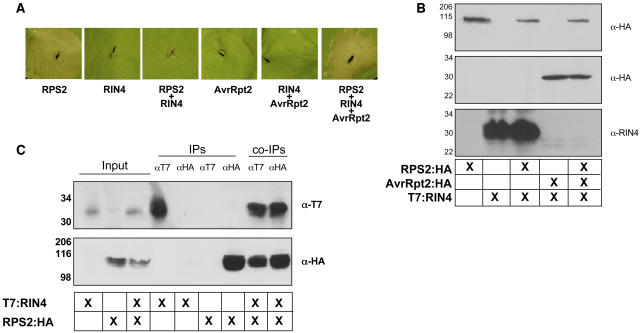
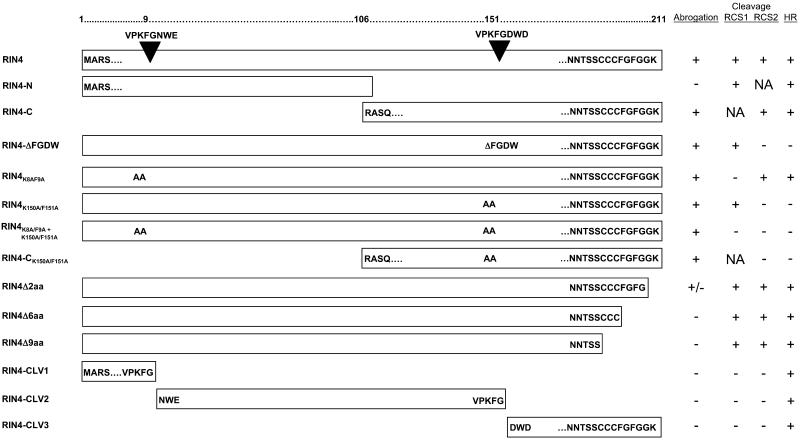
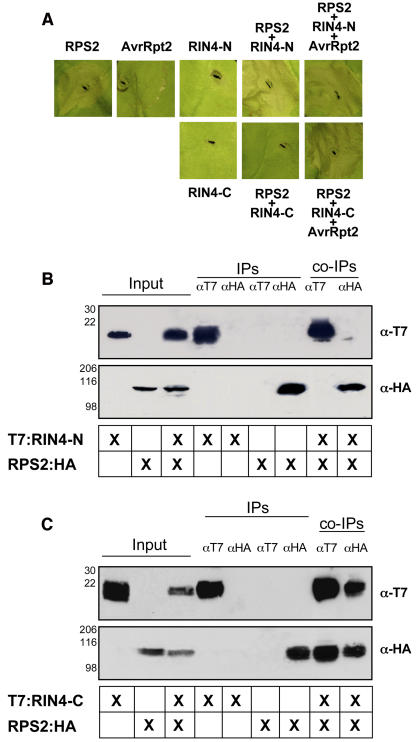
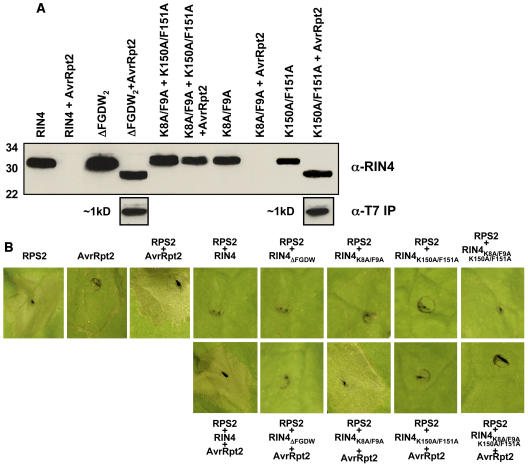
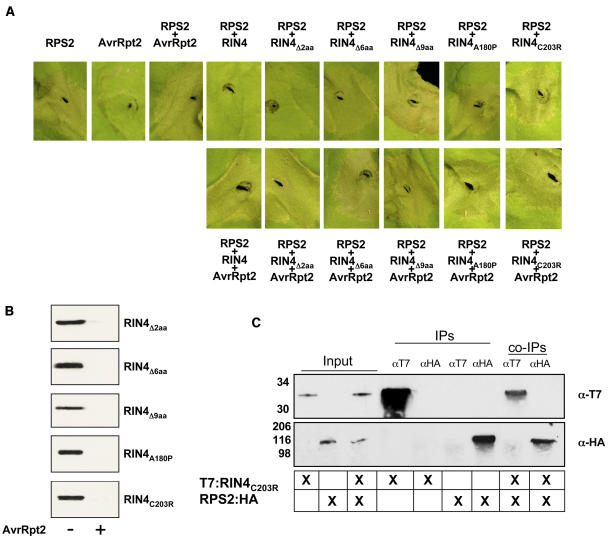
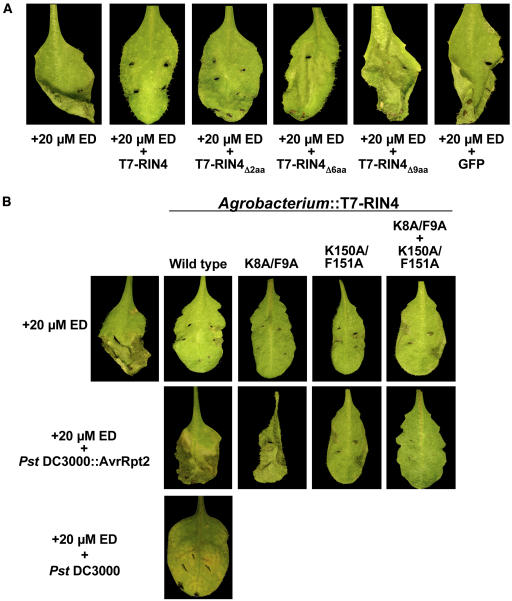
Recent studies have demonstrated that RPS2, a plasma membrane-localized nucleotide binding site/leucine-rich repeat protein from Arabidopsis thaliana, associates with RPM1 Interacting Protein 4 (RIN4) and that this association functions to modulate the RPS2-mediated defense pathway in response to the bacterial effector protein AvrRpt2. In addition to negatively regulating RPS2 activity, RIN4 is also a target of AvrRpt2, a Cys protease and cognate bacterial effector protein of RPS2. Nicotiana benthamiana has been employed as a heterologous expression system to characterize the RPS2-RIN4 association, defining the domains in RIN4 required for the negative regulation of RPS2 activity. Upon inoculation of N. benthamiana leaves with Agrobacterium tumefaciens expressing RPS2, a rapid hypersensitive response (HR) is detected with 22 h of infiltration. The HR can be blocked by infiltrating the leaf with A. tumefaciens expressing RPS2 in the presence of RIN4, recapitulating the ability of RIN4 to interfere with RPS2-mediated resistance in Arabidopsis. Moreover, in the presence of RIN4, the RPS2-mediated HR can be restored by the delivery of AvrRpt2 via A. tumefaciens. This assay has been developed as a phenotypic marker for (1) the HR-inducing phenotype associated with RPS2, (2) negative regulation of RPS2 by RIN4, and (3) the AvrRpt2-mediated disappearance of RIN4. Here, we present a series of deletion and site-directed mutation analyses to identify amino acids in RIN4 required for the RPS2-RIN4 association and to distinguish these from residues in RIN4 that serve as a target sequence for AvrRpt2. In addition to characterizing the RPS2-RIN4 association in N. benthamiana, we have moved forward to show that the biological relevance of these amino acid changes is applicable in Arabidopsis as well. To this end, we have identified specific amino acids within the C-terminal half of RIN4 that are required for RPS2 regulation and association.
Figures
References
-
- Axtell, M.J., Chisholm, S.T., Dahlbeck, D., and Staskawicz, B.J. (2003). Genetic and molecular evidence that the Pseudomonas syringae type III effector protein AvrRpt2 is a cysteine protease. Mol. Microbiol. 49, 1537–1546. - PubMed
-
- Axtell, M.J., and Staskawicz, B.J. (2003). Initiation of RPS2-specified disease resistance in Arabidopsis is coupled to the AvrRpt2-directed elimination of RIN4. Cell 112, 369–377. - PubMed
-
- Belkhadir, Y., Nimchuk, Z., Hubert, D.A., Mackey, D., and Dangl, J.L. (2004. a). Arabidopsis RIN4 negatively regulates disease resistance mediated by RPS2 and RPM1 downstream or independent of the NDR1 signal modulator and is not required for the virulence functions of bacterial type III effectors AvrRpt2 or AvrRpm1. Plant Cell 16, 2822–2835. - PMC - PubMed
-
- Belkhadir, Y., Subramaniam, R., and Dangl, J.L. (2004. b). Plant disease resistance protein signaling: NBS-LRR proteins and their partners. Curr. Opin. Plant Biol. 7, 391–399. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
