

ROCK-I regulates closure of the eyelids and ventral body wall by inducing assembly of actomyosin bundles
- PMID: 15753128
- PMCID: PMC2171774
- DOI: 10.1083/jcb.200411179
ROCK-I regulates closure of the eyelids and ventral body wall by inducing assembly of actomyosin bundles
Abstract
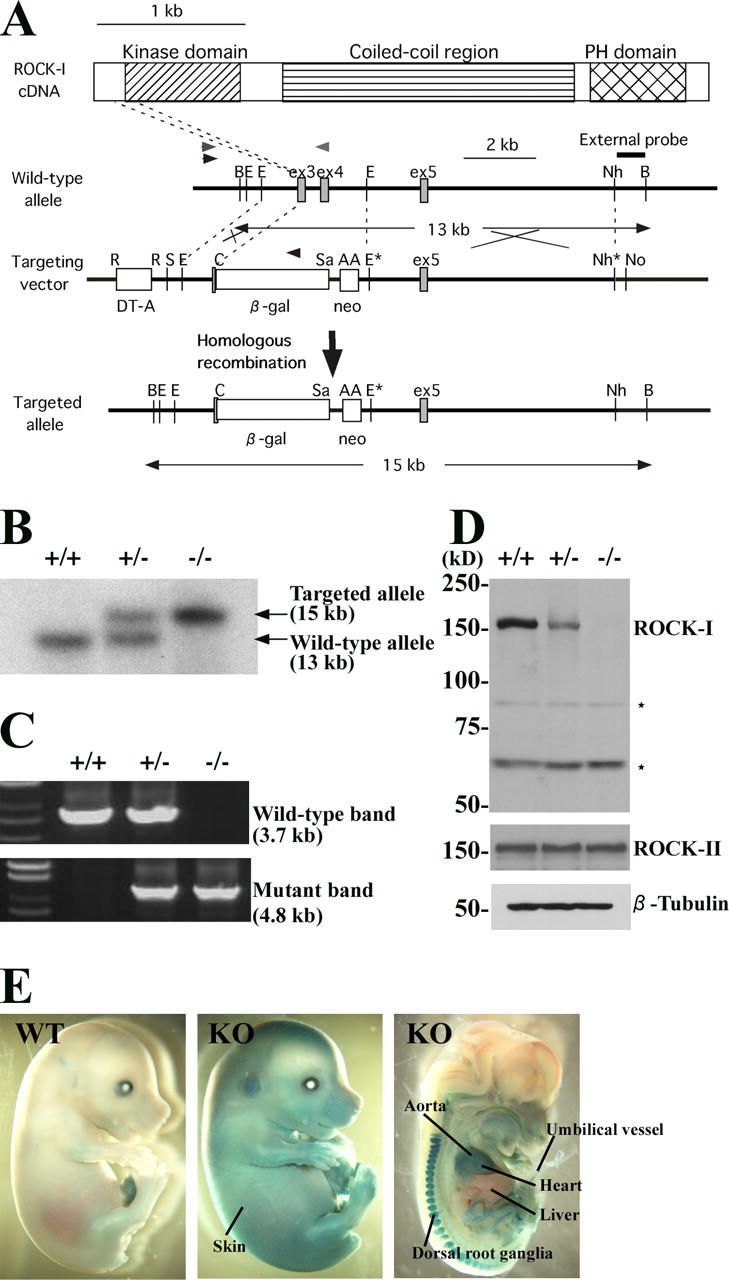
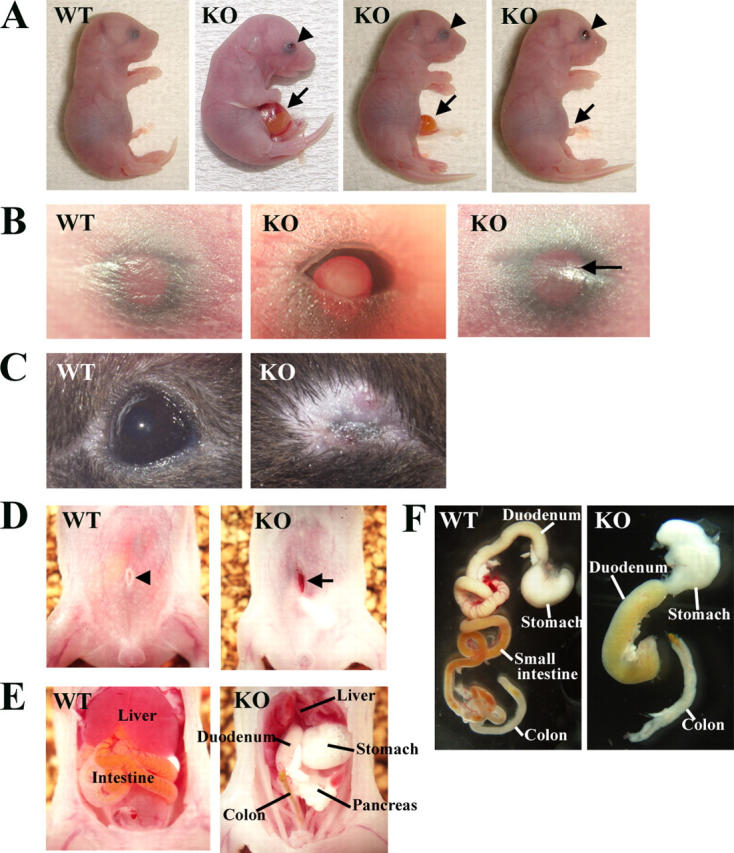
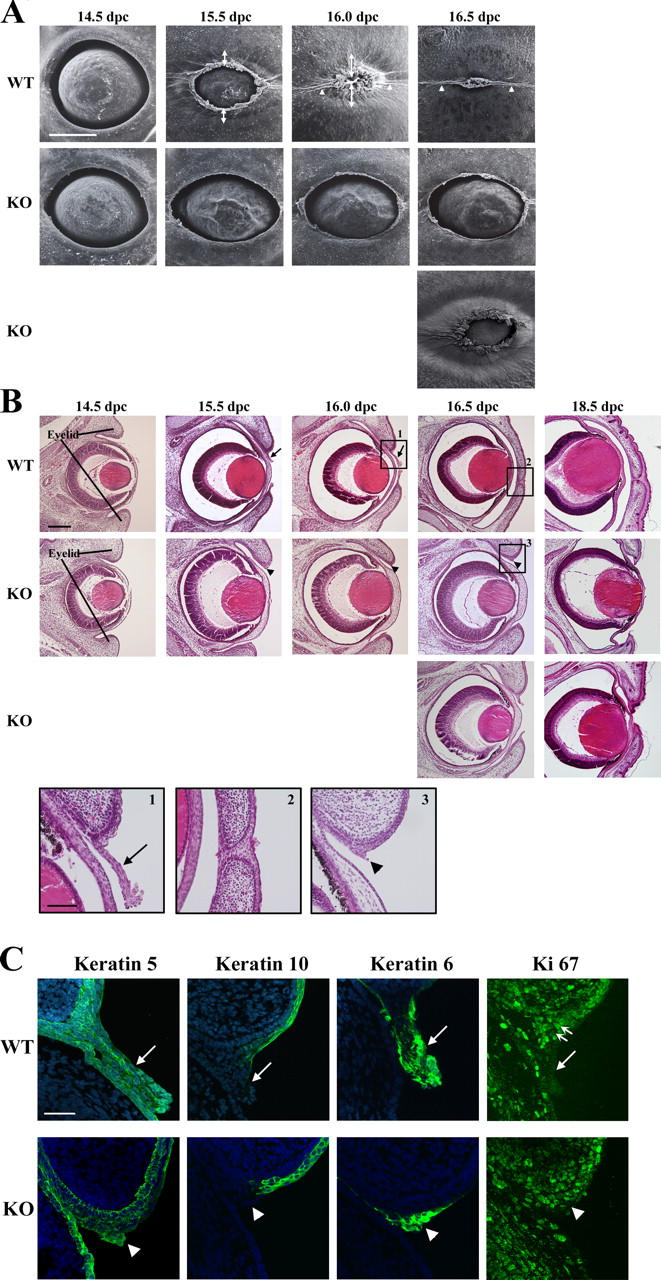
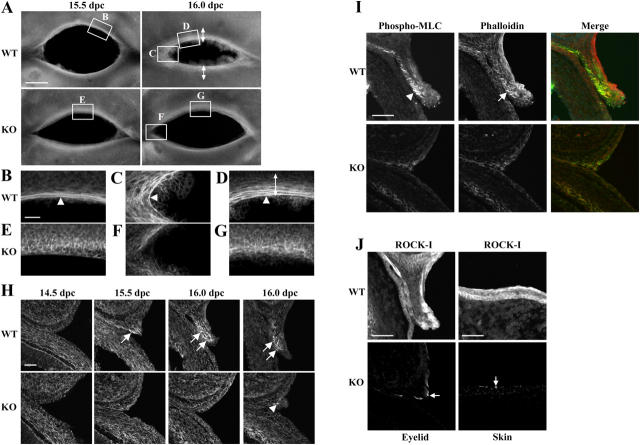
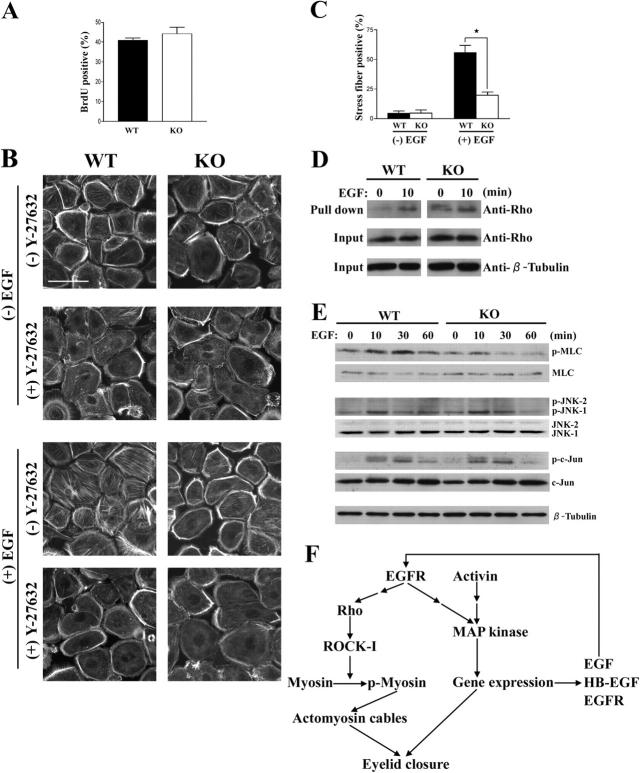
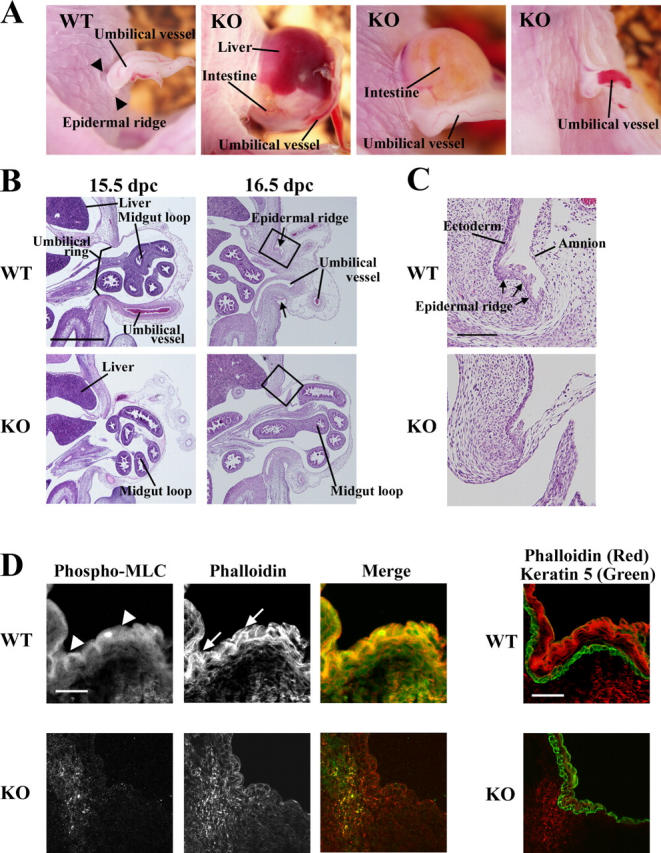
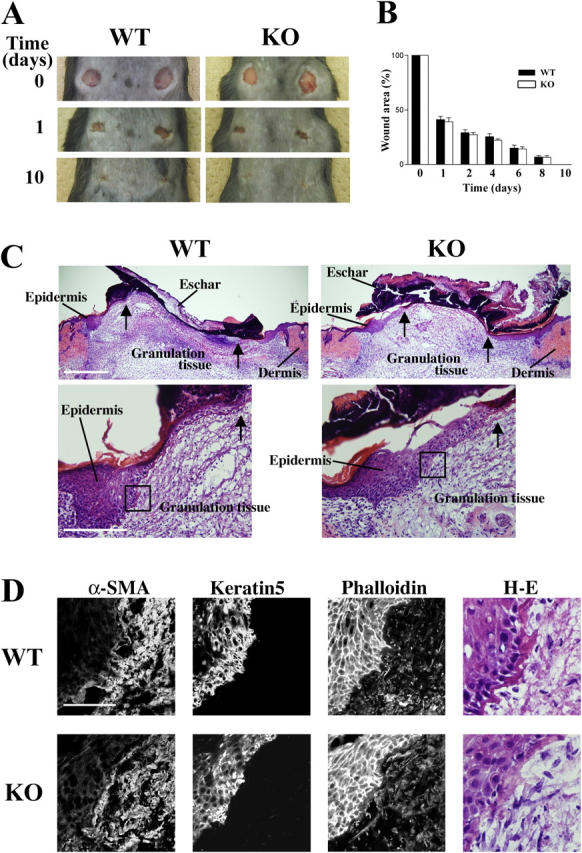
Rho-associated kinase (ROCK) I mediates signaling from Rho to the actin cytoskeleton. To investigate the in vivo functions of ROCK-I, we generated ROCK-I-deficient mice. Loss of ROCK-I resulted in failure of eyelid closure and closure of the ventral body wall, which gave rise to the eyes open at birth and omphalocele phenotypes in neonates. Most ROCK-I(-/-) mice died soon after birth as a result of cannibalization of the omphalocele by the mother. Actin cables that encircle the eye in the epithelial cells of the eyelid were disorganized and accumulation of filamentous actin at the umbilical ring was impaired, with loss of phosphorylation of the myosin regulatory light chain (MLC) at both sites, in ROCK-I(-/-) embryos. Stress fiber formation and MLC phosphorylation induced by EGF were also attenuated in primary keratinocytes from ROCK-I(-/-) mice. These results suggest that ROCK-I regulates closure of the eyelids and ventral body wall through organization of actomyosin bundles.
Figures
References
-
- Amano, M., Y. Fukata, and K. Kaibuchi. 2000. Regulation and functions of Rho-associated kinase. Exp. Cell Res. 261:44–51. - PubMed
-
- Brewer, S., and T. Williams. 2004. Loss of AP-2alpha impacts multiple aspects of ventral body wall development and closure. Dev. Biol. 267:399–417. - PubMed
-
- Etienne-Manneville, S., and A. Hall. 2002. Rho GTPases in cell biology. Nature. 420:629–635. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
