

Interleukin-18 primes the oxidative burst of neutrophils in response to formyl-peptides: role of cytochrome b558 translocation and N-formyl peptide receptor endocytosis
- PMID: 15753257
- PMCID: PMC1065204
- DOI: 10.1128/CDLI.12.3.436-446.2005
Interleukin-18 primes the oxidative burst of neutrophils in response to formyl-peptides: role of cytochrome b558 translocation and N-formyl peptide receptor endocytosis
Abstract
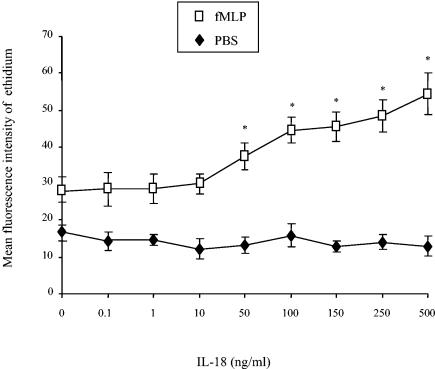
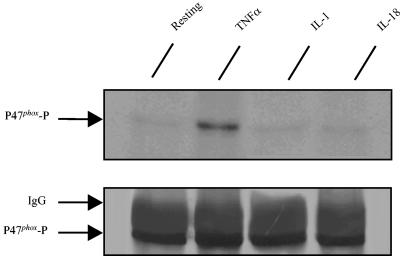
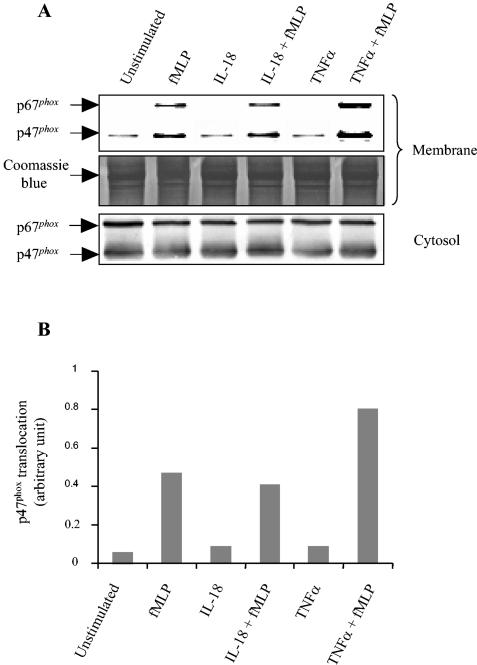
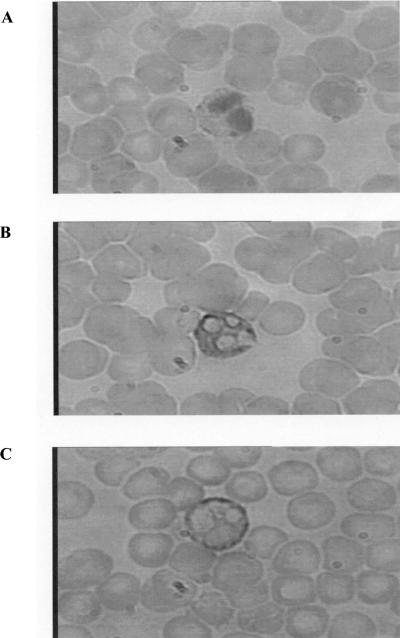
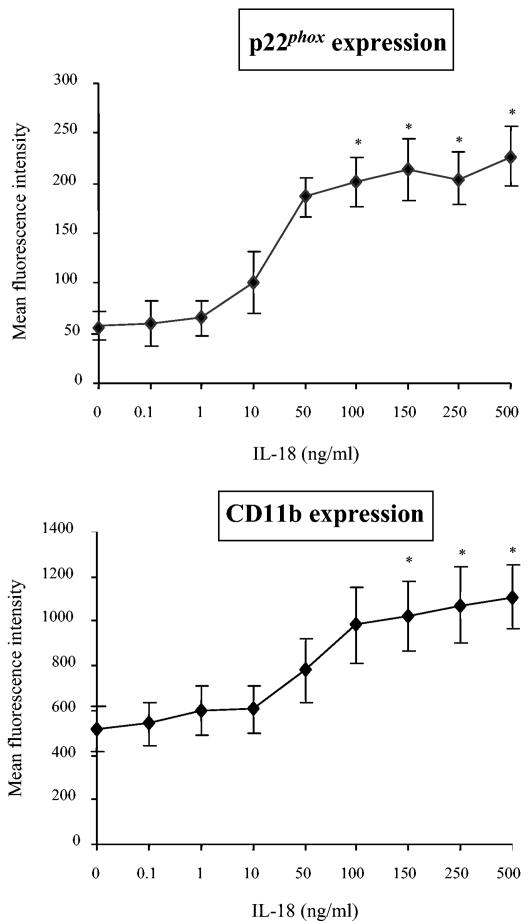
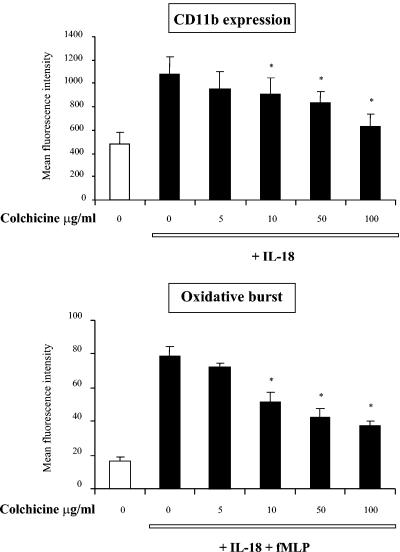
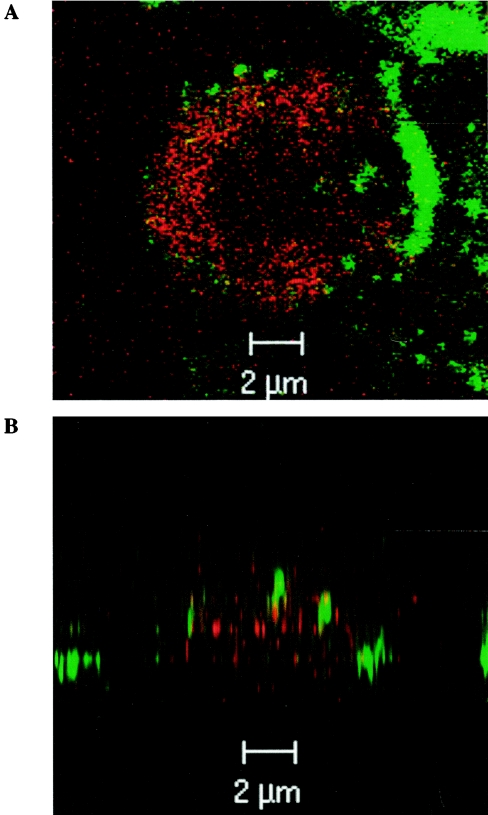
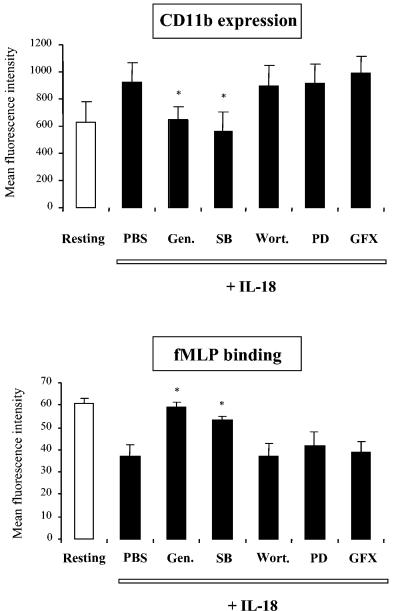
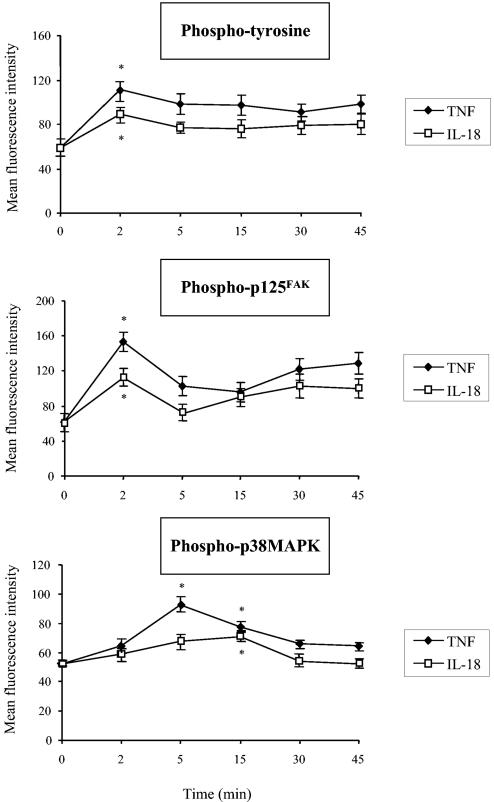
Using flow cytometry, we observed that interleukin-18 (IL-18) primed human neutrophils (PMNs) in whole blood to produce superoxide anion (O2 degrees-) in response to N-formyl peptide (fMLP) stimulation, whereas IL-18 alone had no significant effect. In contrast to tumor necrosis factor alpha (TNF-alpha), which is a cytokine known to strongly prime O2 degrees- production, IL-18 did not induce either p47phox phosphorylation or its translocation from the cytosol to the plasma membrane. However, IL-18 increased PMN degranulation, as shown by increased levels of cytochrome b558 and CD11b expression at the PMN surface. Moreover, addition of IL-18 to whole blood for 45 min reduced the ability of PMNs to bind to fMLP, suggesting endocytosis of fMLP receptors, as visualized by confocal microscopy. 2,3-Butanedione 2-monoxime, which inhibits endosomal recycling of plasma membrane components back to the cell surface, concomitantly accentuated the diminution of fMLP binding at the PMN surface and increased IL-18 priming of O2 degrees- production by PMNs in response to fMLP. This suggests that fMLP receptor endocytosis could account, at least in part, for the priming of O2 degrees- production. In addition, genistein, a tyrosine kinase inhibitor, and SB203580, a p38 mitogen-activated protein kinase (p38MAPK) inhibitor, completely reversed the decreased level of fMLP binding and increased the level of CD11b expression after IL-18 treatment. Flow cytometric analysis of intact PMNs in whole blood showed that IL-18 increased p38MAPK phosphorylation and tyrosine phosphorylation. In particular, IL-18 induced phosphorylation of focal adhesion kinase (p125FAK), which has been implicated in cytoskeleton reorganization. Taken together, our findings suggest several mechanisms that are likely to regulate cytokine-induced priming of the oxidative burst in PMNs in their blood environment.
Figures
Similar articles
-
Stimulation of human neutrophils with formyl-methionyl-leucyl-phenylalanine induces tyrosine phosphorylation and activation of two distinct mitogen-activated protein-kinases.J Immunol. 1993 Feb 15;150(4):1563-77. J Immunol. 1993. PMID: 7679431
-
[Modulation of the oxidative burst of human neutrophils by pro- and anti-inflammatory cytokines].Pathol Biol (Paris). 1996 Jan;44(1):36-41. Pathol Biol (Paris). 1996. PMID: 8734298 Review. French.
-
5-Hydroxy-7-methoxyflavone inhibits N-formyl-L-methionyl-L-leucyl-L-phenylalanine-induced superoxide anion production by specific modulate membrane localization of Tec with a PI3K independent mechanism in human neutrophils.Biochem Pharmacol. 2012 Jul 15;84(2):182-91. doi: 10.1016/j.bcp.2012.03.015. Epub 2012 Mar 29. Biochem Pharmacol. 2012. PMID: 22484311
-
Local anesthetics inhibit priming of neutrophils by lipopolysaccharide for enhanced release of superoxide: suppression of cytochrome b558 expression by disparate mechanisms.J Leukoc Biol. 2005 Dec;78(6):1356-65. doi: 10.1189/jlb.0305169. Epub 2005 Oct 4. J Leukoc Biol. 2005. PMID: 16204644
-
[Regulation of human neutrophil oxidative burst by pro- and anti-inflammatory cytokines].J Soc Biol. 2002;196(1):37-46. J Soc Biol. 2002. PMID: 12134631 Review. French.
Cited by
-
IL-17A potentiates TNFα-induced secretion from human endothelial cells and alters barrier functions controlling neutrophils rights of passage.Pflugers Arch. 2014 May;466(5):961-72. doi: 10.1007/s00424-013-1354-5. Epub 2013 Sep 27. Pflugers Arch. 2014. PMID: 24072078 Free PMC article.
-
The role of interleukin-1 and interleukin-18 in pro-inflammatory and anti-viral responses to rhinovirus in primary bronchial epithelial cells.PLoS One. 2013 May 28;8(5):e63365. doi: 10.1371/journal.pone.0063365. Print 2013. PLoS One. 2013. PMID: 23723976 Free PMC article.
-
Exocytosis of neutrophil granule subsets and activation of prolyl isomerase 1 are required for respiratory burst priming.J Innate Immun. 2013;5(3):277-89. doi: 10.1159/000345992. Epub 2013 Jan 26. J Innate Immun. 2013. PMID: 23363774 Free PMC article. Clinical Trial.
-
Inflammatory cytokines are associated to lower glomerular filtration rate in patients with hypertensive crisis.Front Cardiovasc Med. 2022 Sep 29;9:969339. doi: 10.3389/fcvm.2022.969339. eCollection 2022. Front Cardiovasc Med. 2022. PMID: 36247461 Free PMC article.
-
Acute COVID-19 and LongCOVID syndrome - molecular implications for therapeutic strategies - review.Front Immunol. 2025 Apr 17;16:1582783. doi: 10.3389/fimmu.2025.1582783. eCollection 2025. Front Immunol. 2025. PMID: 40313948 Free PMC article. Review.
References
-
- Babior, B. M. 1984. Oxidants from phagocytes: agents of defense and destruction. Blood 64:959-966. - PubMed
-
- Babior, B. M. 1999. NADPH oxidase: an update. Blood 93:1464-1476. - PubMed
-
- Barlic, J., J. D. Andrews, A. A. Kelvin, S. E. Bosinger, M. E. DeVries, L. Xu, T. Dobransky, R. D. Feldman, S. S. Ferguson, and D. J. Kelvin. 2000. Regulation of tyrosine kinase activation and granule release through β-arrestin by CXCR1. Nat. Immunol. 87:227-233. - PubMed
-
- Chanock, S. J., J. El Benna, R. M. Smith, and B. M. Babior. 1994. The respiratory burst oxidase. J. Biol. Chem. 269:24519-24522. - PubMed
-
- Coelho, A. L. J., M. S. De Freitas, A. Mariano-Oliveira, A. L. Oliveira-Carvalho, R. B. Zingali, and C. Barja-Fidalgo. 2001. Interaction of disintegrins with human neutrophils induces cytoskeleton reorganization, focal adhesion kinase activation, and extracellular-regulated kinase-2 nuclear translocation, interfering with the chemotactic function. FASEB J. 15:1643-1645. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
