

Developmentally regulated instability of the GPI-PLC mRNA is dependent on a short-lived protein factor
- PMID: 15755751
- PMCID: PMC1062878
- DOI: 10.1093/nar/gki298
Developmentally regulated instability of the GPI-PLC mRNA is dependent on a short-lived protein factor
Abstract
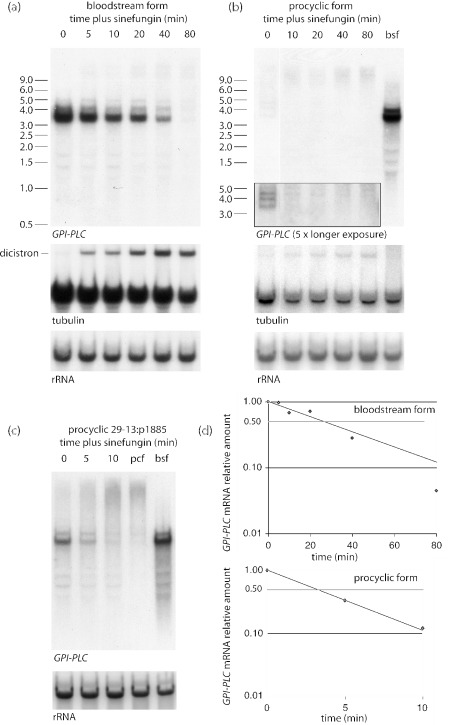
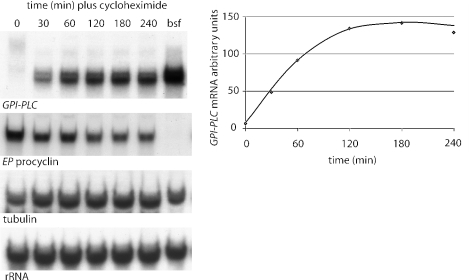
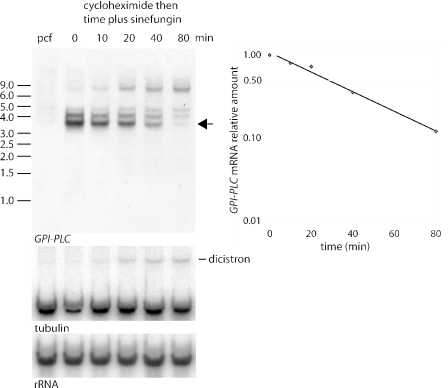
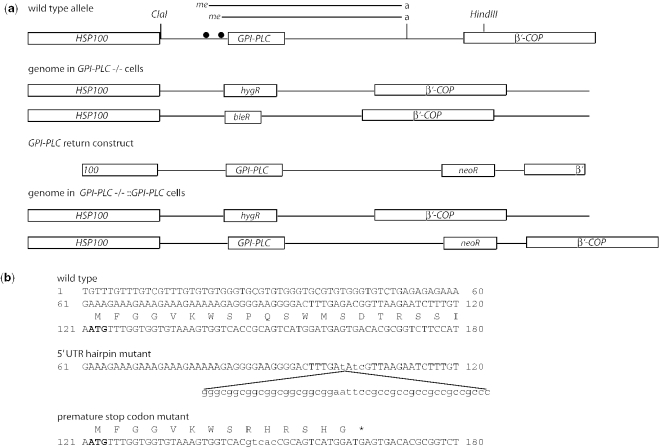
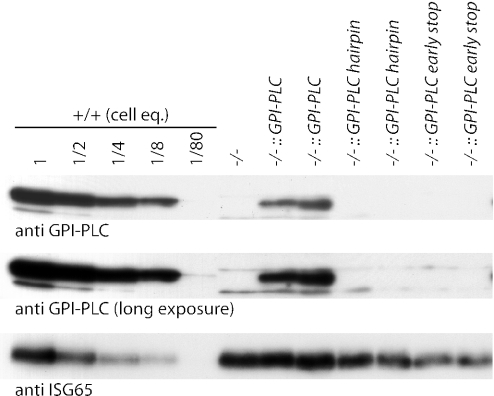
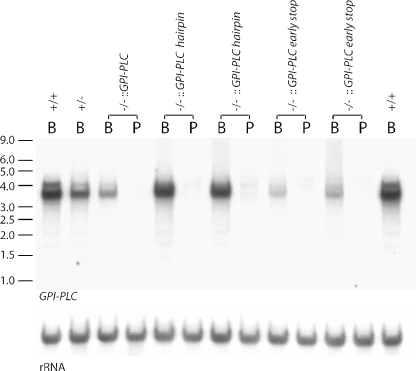
The expression of the vast majority of protein coding genes in trypanosomes is regulated exclusively at the post-transcriptional level. Developmentally regulated mRNAs that vary in levels of expression have provided an insight into one mechanism of regulation; a decrease in abundance is due to a shortened mRNA half-life. The decrease in half-life involves cis-acting elements in the 3' untranslated region of the mRNA. The trans-acting factors necessary for the increased rate of degradation remain uncharacterized. The GPI-PLC gene in Trypanosoma brucei encodes a phospholipase C expressed in mammalian bloodstream form, but not in the insect procyclic form. Here, it is reported that the differential expression of the GPI-PLC mRNA also results from a 10-fold difference in half-life. Second, the instability of the GPI-PLC mRNA in procyclic forms can be reversed by the inhibition of protein synthesis. Third, specifically blocking the translation of the GPI-PLC mRNA in procyclic forms by the inclusion of a hairpin in the 5' untranslated region does not result in stabilization of the mRNA. Thus, the effect of protein synthesis inhibitors in stabilizing the GPI-PLC mRNA operates in trans through a short-lived factor dependent on protein synthesis.
Figures
Similar articles
-
A function for a specific zinc metalloprotease of African trypanosomes.PLoS Pathog. 2007 Oct 19;3(10):1432-45. doi: 10.1371/journal.ppat.0030150. PLoS Pathog. 2007. PMID: 17953481 Free PMC article.
-
A novel strategy to identify the location of necessary and sufficient cis-acting regulatory mRNA elements in trypanosomes.RNA. 2005 Jul;11(7):1108-16. doi: 10.1261/rna.2510505. Epub 2005 May 31. RNA. 2005. PMID: 15928343 Free PMC article.
-
Regulation of surface coat exchange by differentiating African trypanosomes.Mol Biochem Parasitol. 2006 Jun;147(2):211-23. doi: 10.1016/j.molbiopara.2006.02.013. Epub 2006 Mar 9. Mol Biochem Parasitol. 2006. PMID: 16564583
-
The properties and function of the glycosylphosphatidylinositol-phospholipase C in Trypanosoma brucei.Mol Biochem Parasitol. 1998 Mar 1;91(1):153-64. doi: 10.1016/s0166-6851(97)00190-4. Mol Biochem Parasitol. 1998. PMID: 9574933 Review.
-
RNA-binding proteins and mRNA turnover in trypanosomes.Trends Parasitol. 2003 Apr;19(4):151-5. doi: 10.1016/s1471-4922(03)00035-7. Trends Parasitol. 2003. PMID: 12689640 Review.
Cited by
-
LiZIP3 is a cellular zinc transporter that mediates the tightly regulated import of zinc in Leishmania infantum parasites.Mol Microbiol. 2015 May;96(3):581-95. doi: 10.1111/mmi.12957. Epub 2015 Mar 11. Mol Microbiol. 2015. PMID: 25644708 Free PMC article.
-
Proteome remodelling during development from blood to insect-form Trypanosoma brucei quantified by SILAC and mass spectrometry.BMC Genomics. 2012 Oct 16;13:556. doi: 10.1186/1471-2164-13-556. BMC Genomics. 2012. PMID: 23067041 Free PMC article.
-
The cell biology of Trypanosoma brucei differentiation.Curr Opin Microbiol. 2007 Dec;10(6):539-46. doi: 10.1016/j.mib.2007.09.014. Epub 2007 Nov 9. Curr Opin Microbiol. 2007. PMID: 17997129 Free PMC article. Review.
-
A high-throughput protein tagging toolkit that retains endogenous untranslated regions for studying gene regulation in kinetoplastids.Open Biol. 2025 Feb;15(2):240334. doi: 10.1098/rsob.240334. Epub 2025 Feb 26. Open Biol. 2025. PMID: 39999874 Free PMC article.
-
Simultaneous detection of mRNA transcription and decay intermediates by dual colour single mRNA FISH on subcellular resolution.Nucleic Acids Res. 2017 Apr 20;45(7):e49. doi: 10.1093/nar/gkw1245. Nucleic Acids Res. 2017. PMID: 27940558 Free PMC article.
References
-
- Hall N., Berriman M., Lennard N.J., Harris B.R., Hertz-Fowler C., Bart-Delabesse E.N., Gerrard C.S., Atkin R.J., Barron A.J., Bowman S., et al. The DNA sequence of chromosome I of an African trypanosome: gene content, chromosome organisation, recombination and polymorphism. Nucleic Acids Res. 2003;31:4864–4873. - PMC - PubMed
-
- Kooter J.M., Van der Spek H.J., Wagter R., d'Oliviera C.E., Van der Hoeven F., Johnson P.J., Borst P. The anatomy and transcription of a telomeric expression site for variant specific surface antigens in Trypanosoma brucei. Cell. 1987;51:261–272. - PubMed
-
- Johnson P.J., Kooter J.M., Borst P. Inactivation of transcription by UV irradiation of T. brucei provides evidence for a multicistronic transcription unit including a VSG gene. Cell. 1987;51:273–281. - PubMed
