

Agonist activation of arachidonate-regulated Ca2+-selective (ARC) channels in murine parotid and pancreatic acinar cells
- PMID: 15760932
- PMCID: PMC1464460
- DOI: 10.1113/jphysiol.2005.085704
Agonist activation of arachidonate-regulated Ca2+-selective (ARC) channels in murine parotid and pancreatic acinar cells
Abstract
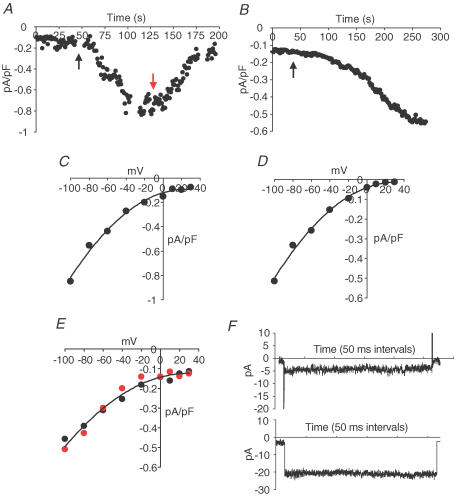
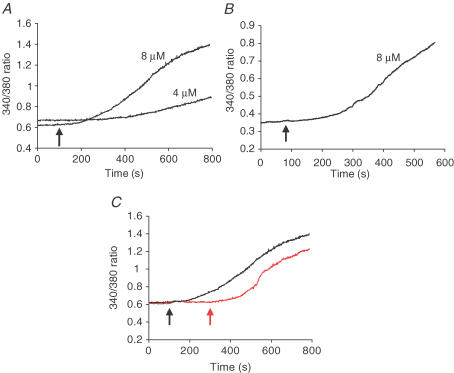
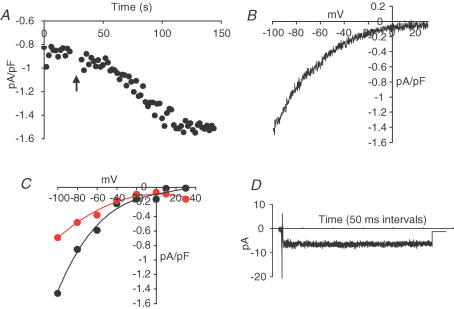
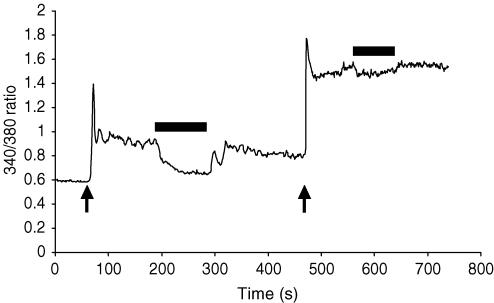
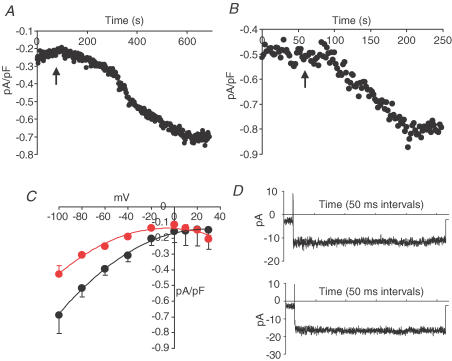
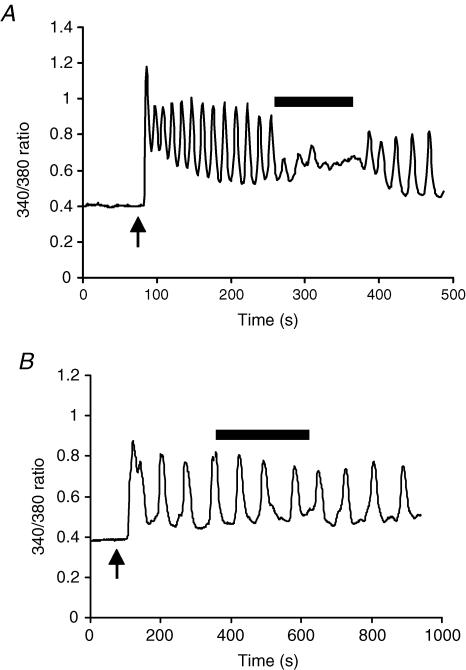
ARC channels (arachidonate-regulated Ca(2+)-selective channels) are a novel type of highly Ca(2+)-selective channel that are specifically activated by low concentrations of agonist-induced arachidonic acid. This activation occurs in the absence of any depletion of internal Ca(2+) stores (i.e. they are 'non-capacitative'). Previous studies in HEK293 cells have shown that these channels provide the predominant pathway for the entry of Ca(2+) seen at low agonist concentrations where oscillatory [Ca(2+)](i) signals are typically produced. In contrast, activation of the more widely studied store-operated Ca(2+) channels (e.g. CRAC channels) is only seen at higher agonist concentrations where sustained 'plateau-type'[Ca(2+)](i) responses are observed. We have now demonstrated the presence of ARC channels in both parotid and pancreatic acinar cells and shown that, again, they are specifically activated by the low concentrations of appropriate agonists (carbachol in the parotid, and both carbachol and cholecystokinin in the pancreas) that are associated with oscillatory [Ca(2+)](i) signals in these cells. Uncoupling the receptor-mediated activation of cytosolic phospholipase A(2) (cPLA(2)) with isotetrandrine reduces the activation of the ARC channels by carbachol and, correspondingly, markedly inhibits the [Ca(2+)](i) signals induced by low carbachol concentrations, whilst those signals seen at high agonist concentrations are essentially unaffected. Interestingly, in the pancreatic acinar cells, activation by cholecystokinin induces a current through the ARC channels that is only approximately 60% of that seen with carbachol. This is consistent with previous reports indicating that carbachol-induced [Ca(2+)](i) signals in these cells are much more dependent on Ca(2+) entry than are the cholecystokinin-induced responses.
Figures
References
-
- Begenisich T, Nakamoto T, Ovitt CE, Nehrke K, Brugnara C, Alper SL, Melvin JE. Physiological roles of the intermediate conductance, Ca2+-activated potassium channel Kcnn4. J Biol Chem. 2004;279:47681–47687. - PubMed
-
- Bers DM, Patton CW, Nuccitelli R. A practical guide to the preparation of Ca2+ buffers. Meths Cell Biol. 1994;40:1–29. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous