

Creating a sense of auditory space
- PMID: 15760940
- PMCID: PMC1464715
- DOI: 10.1113/jphysiol.2005.083113
Creating a sense of auditory space
Abstract
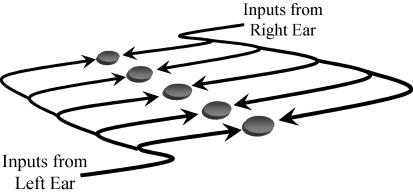
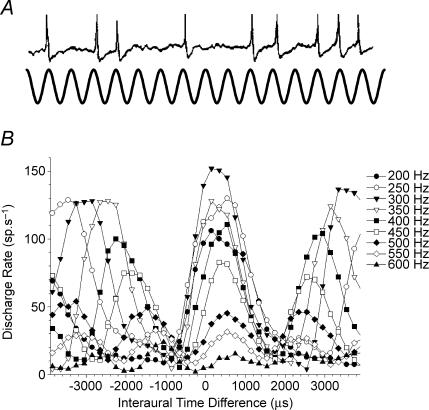
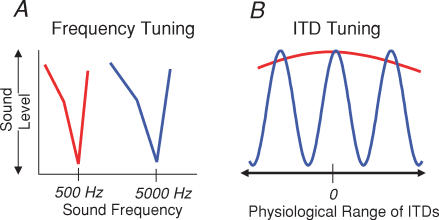
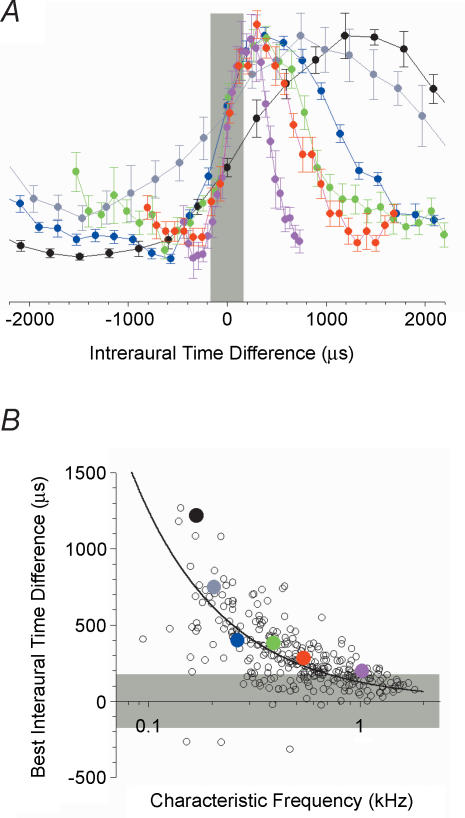
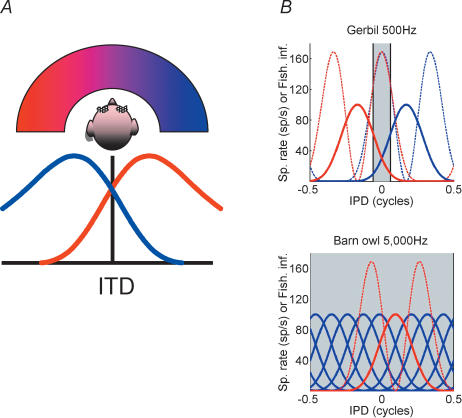
Determining the location of a sound source requires the use of binaural hearing--information about a sound at the two ears converges onto neurones in the auditory brainstem to create a binaural representation. The main binaural cue used by many mammals to locate a sound source is the interaural time difference, or ITD. For over 50 years a single model has dominated thinking on how ITDs are processed. The Jeffress model consists of an array of coincidence detectors--binaural neurones that respond maximally to simultaneous input from each ear--innervated by a series of delay lines--axons of varying length from the two ears. The purpose of this arrangement is to create a topographic map of ITD, and hence spatial position in the horizontal plane, from the relative timing of a sound at the two ears. This model appears to be realized in the brain of the barn owl, an auditory specialist, and has been assumed to hold for mammals also. Recent investigations, however, indicate that both the means by which neural tuning for preferred ITD, and the coding strategy used by mammals to determine the location of a sound source, may be very different to barn owls and to the model proposed by Jeffress.
Figures
Comment in
-
The senses.J Physiol. 2005 Jul 1;566(Pt 1):5. doi: 10.1113/jphysiol.2005.090837. Epub 2005 May 26. J Physiol. 2005. PMID: 15919707 Free PMC article. No abstract available.
References
-
- Adams JC, Warr WB. Origins of axons in the cat's acoustic striae determined by injection of horseradish peroxidase into severed tracts. J Comp Neurol. 1976;170:107–121. - PubMed
-
- Bala AD, Spitzer MW, Takahashi TT. Prediction of auditory spatial acuity from neural images on the owl's auditory space map. Nature. 2003;424:771–774. - PubMed
-
- Brand A, Behrend O, Marquardt T, McAlpine D, Grothe B. Precise inhibition is essential for microsecond interaural time difference coding. Nature. 2002;417:543–547. - PubMed
-
- Breebaart J, van de Par S, Kohlrausch A. Binaural processing model based on contralateral inhibition. I. Model structure. J Acoust Soc Am. 2001;110:1074–1088. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources