

Calcium oscillations in interstitial cells of the rabbit urethra
- PMID: 15760947
- PMCID: PMC1464513
- DOI: 10.1113/jphysiol.2004.078097
Calcium oscillations in interstitial cells of the rabbit urethra
Abstract
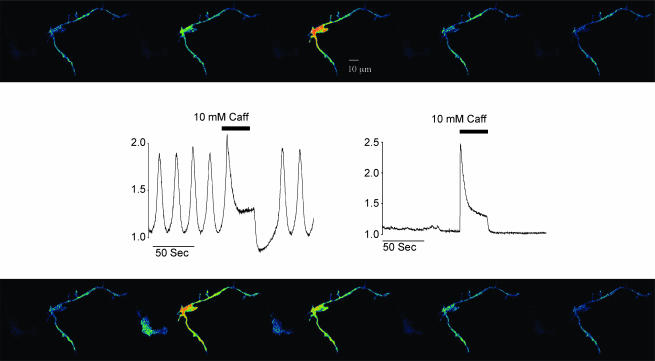
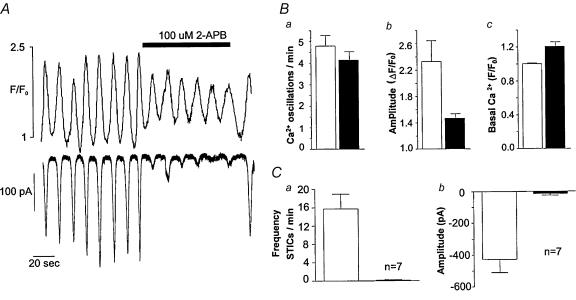
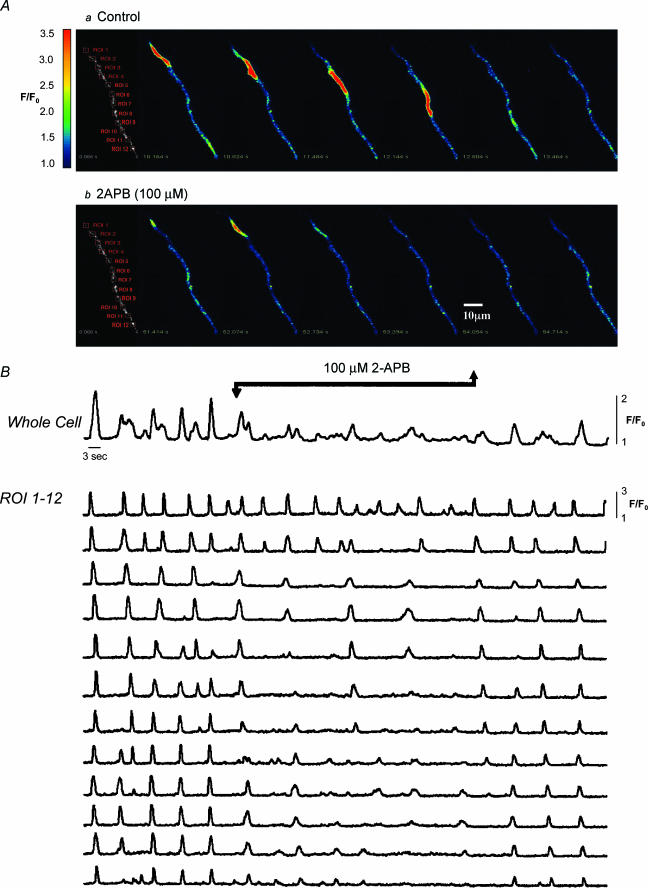
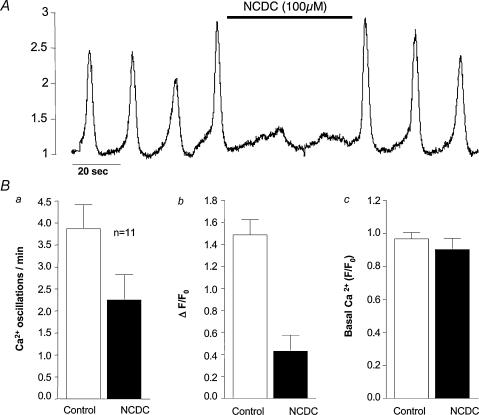
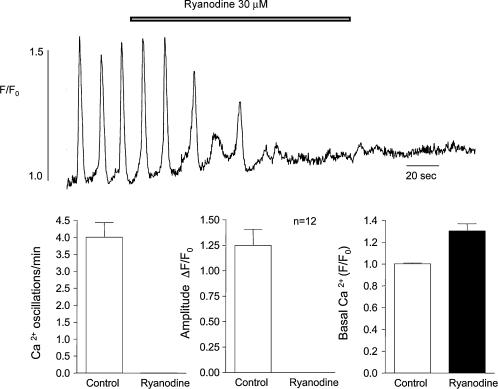
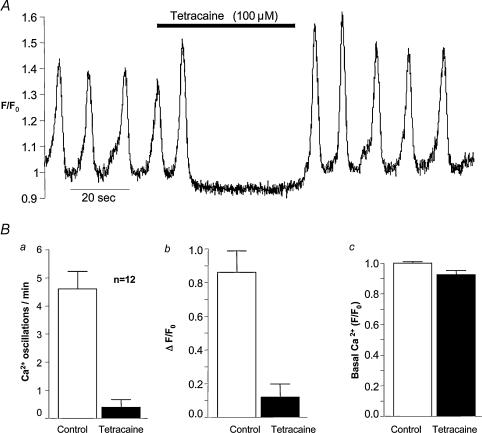
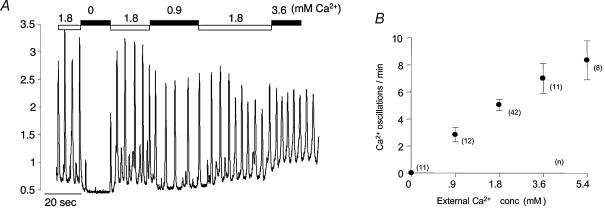
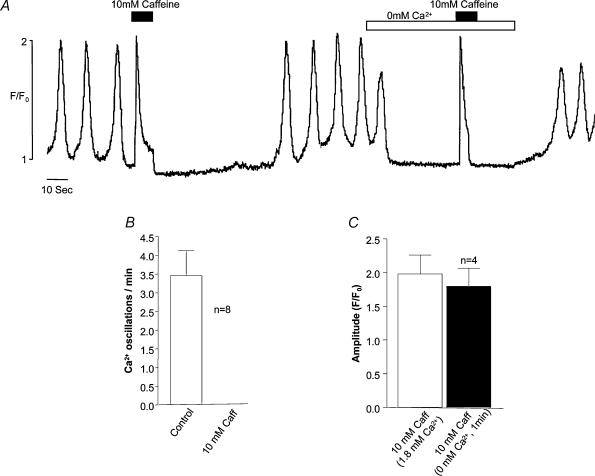
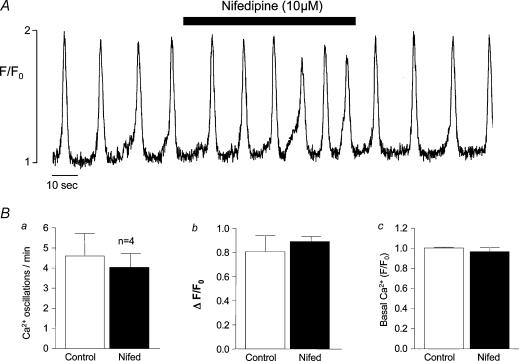
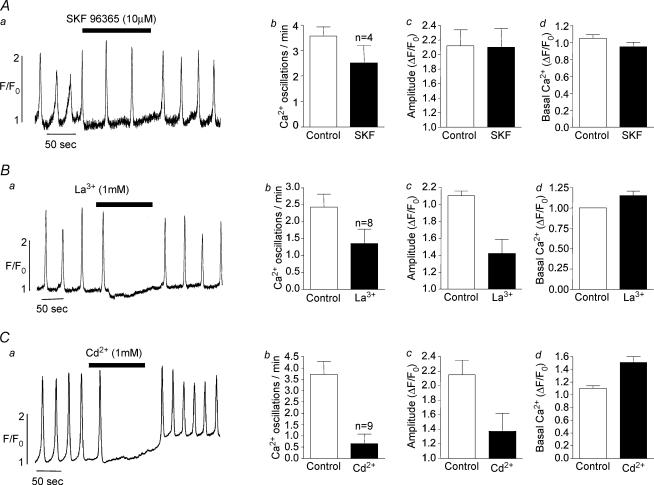
Measurements were made (using fast confocal microscopy) of intracellular Ca2+ levels in fluo-4 loaded interstitial cells isolated from the rabbit urethra. These cells exhibited regular Ca2+ oscillations which were associated with spontaneous transient inward currents recorded under voltage clamp. Interference with D-myo-inositol 1,4,5-trisphosphate (IP3) induced Ca2+ release using 100 microm 2-aminoethoxydiphenyl borate, and the phospholipase C (PLC) inhibitors 2-nitro-4-carboxyphenyl N,N-diphenylcarbamate and U73122 decreased the amplitude of spontaneous oscillations but did not abolish them. However, oscillations were abolished when ryanodine receptors were blocked with tetracaine or ryanodine. Oscillations ceased in the absence of external Ca2+, and frequency was directly proportional to the external Ca2+ concentration. Frequency of Ca2+ oscillation was reduced by SKF-96365, but not by nifedipine. Lanthanum and cadmium completely blocked oscillations. These results suggest that Ca2+ oscillations in isolated rabbit urethral interstitial cells are initiated by Ca2+ release from ryanodine-sensitive intracellular stores, that oscillation frequency is very sensitive to the external Ca2+ concentration and that conversion of the primary oscillation to a propagated Ca2+ wave depends upon IP3-induced Ca2+ release.
Figures
References
-
- Bayguinov O, Hagen B, Bonev AD, Nelson MT, Sanders KM. Intracellular calcium events activated by ATP in murine colonic myocytes. Am J Physiol Cell Physiol. 2000;279:C126–135. - PubMed
-
- Boittin FX, Coussin F, Macrez N, Mironneau C, Mironneau J. Inositol 1,4,5-trisphosphate-and ryanodine-sensitive Ca2+ release channel-dependent Ca2+ signalling in rat portal vein myocytes. Cell Calcium. 1998;23:303–311. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous