

CFTR gating II: Effects of nucleotide binding on the stability of open states
- PMID: 15767296
- PMCID: PMC1289160
- DOI: 10.1085/jgp.200409228
CFTR gating II: Effects of nucleotide binding on the stability of open states
Abstract
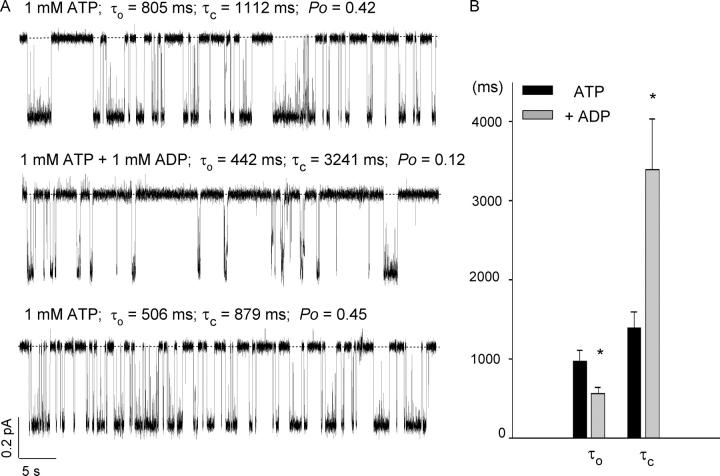
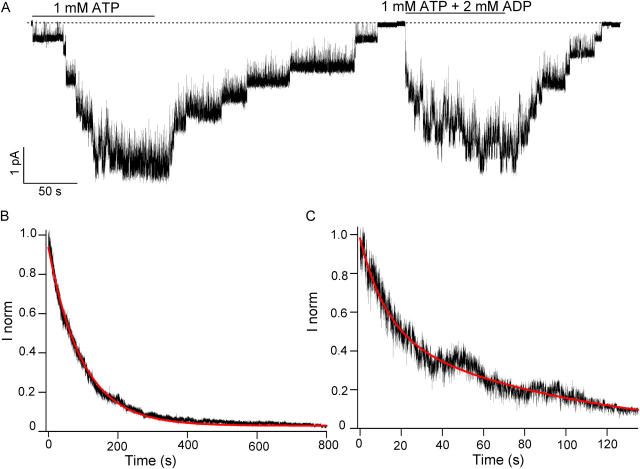
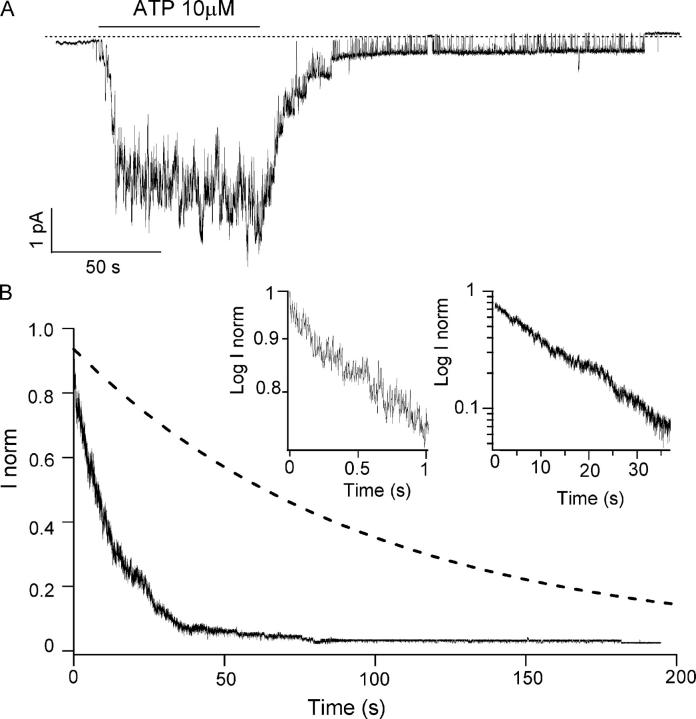
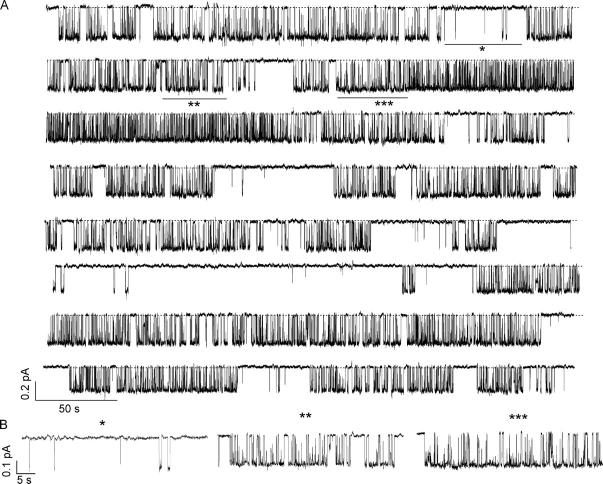
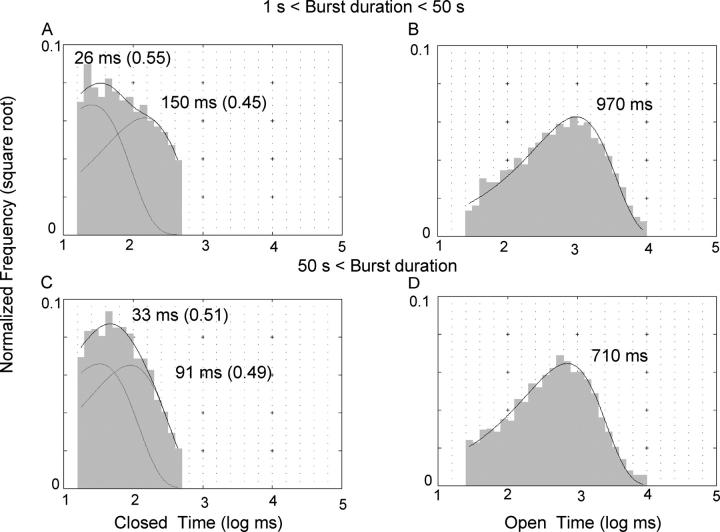
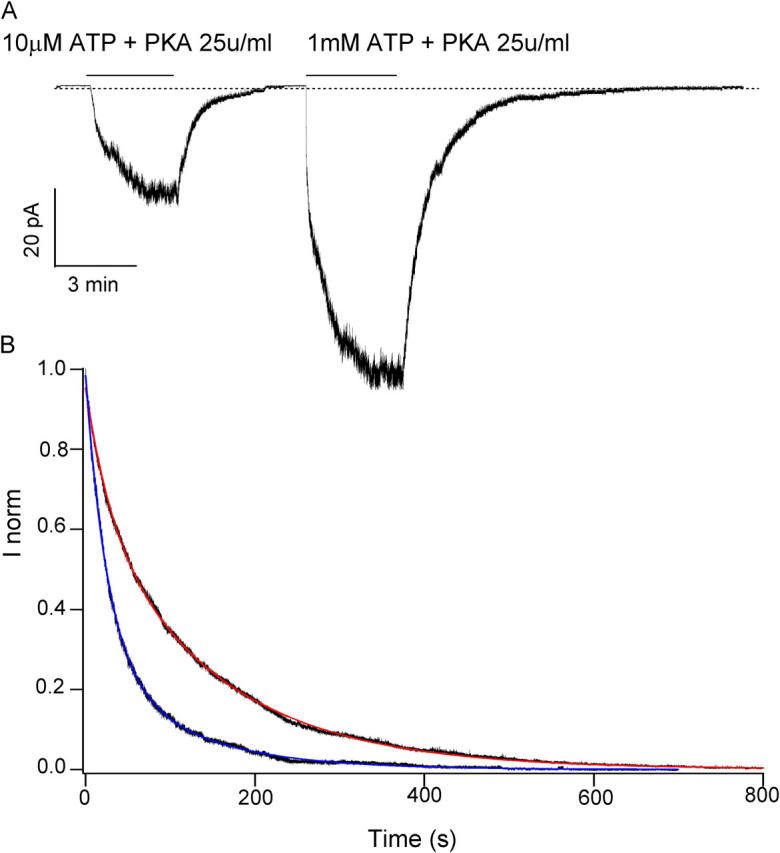
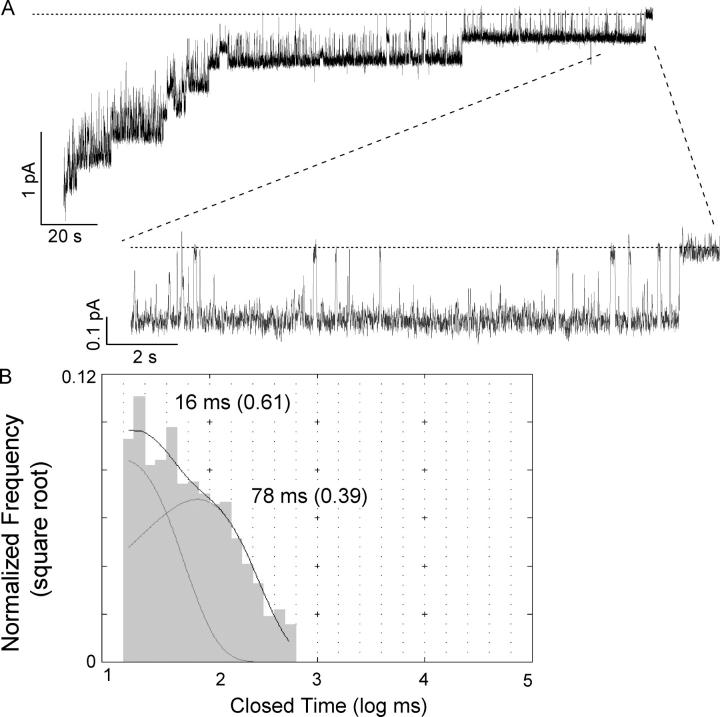
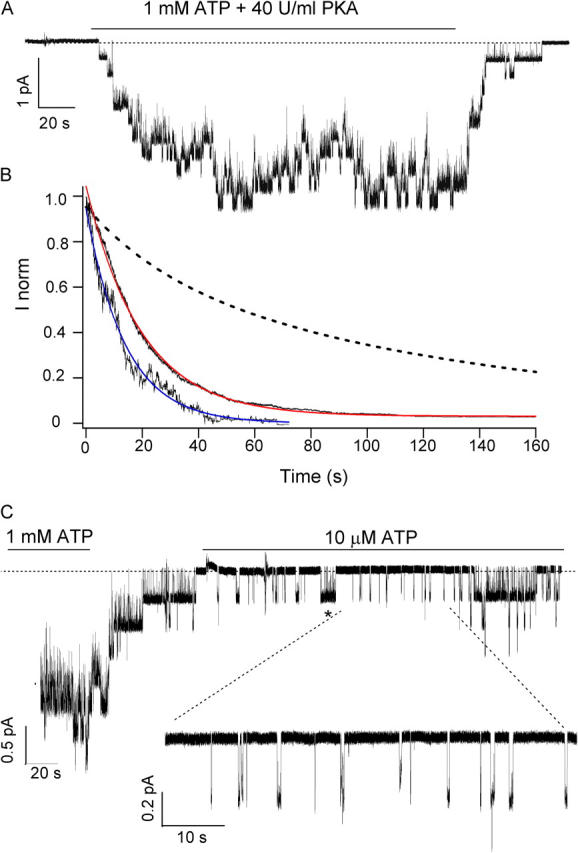
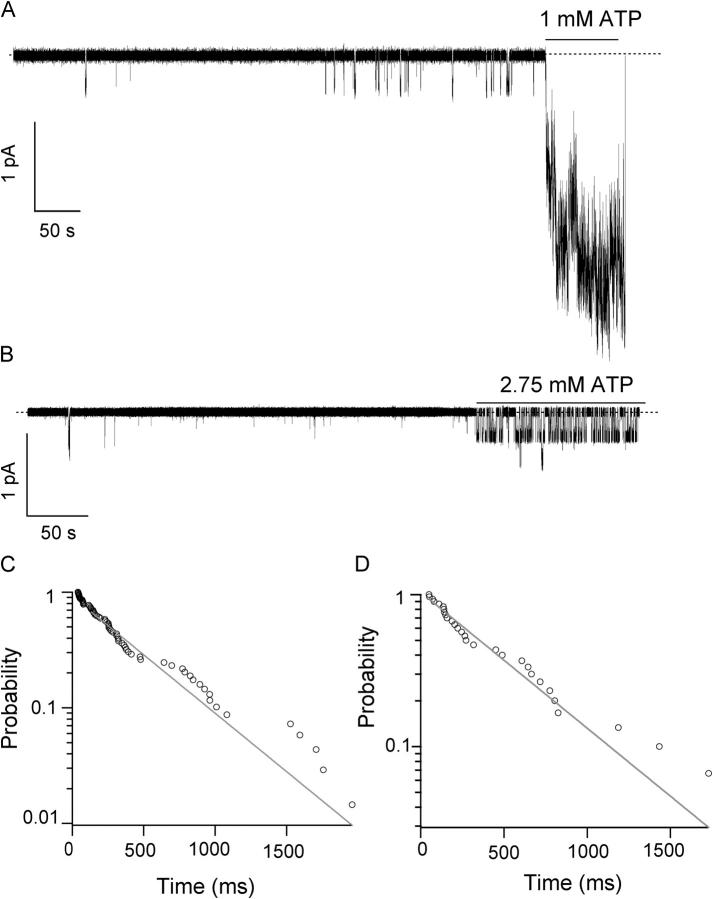
Previously, we demonstrated that ADP inhibits cystic fibrosis transmembrane conductance regulator (CFTR) opening by competing with ATP for a binding site presumably in the COOH-terminal nucleotide binding domain (NBD2). We also found that the open time of the channel is shortened in the presence of ADP. To further study this effect of ADP on the open state, we have used two CFTR mutants (D1370N and E1371S); both have longer open times because of impaired ATP hydrolysis at NBD2. Single-channel kinetic analysis of DeltaR/D1370N-CFTR shows unequivocally that the open time of this mutant channel is decreased by ADP. DeltaR/E1371S-CFTR channels can be locked open by millimolar ATP with a time constant of approximately 100 s, estimated from current relaxation upon nucleotide removal. ADP induces a shorter locked-open state, suggesting that binding of ADP at a second site decreases the locked-open time. To test the functional consequence of the occupancy of this second nucleotide binding site, we changed the [ATP] and performed similar relaxation analysis for E1371S-CFTR channels. Two locked-open time constants can be discerned and the relative distribution of each component is altered by changing [ATP] so that increasing [ATP] shifts the relative distribution to the longer locked-open state. Single-channel kinetic analysis for DeltaR/E1371S-CFTR confirms an [ATP]-dependent shift of the distribution of two locked-open time constants. These results support the idea that occupancy of a second ATP binding site stabilizes the locked-open state. This binding site likely resides in the NH2-terminal nucleotide binding domain (NBD1) because introducing the K464A mutation, which decreases ATP binding affinity at NBD1, into E1371S-CFTR shortens the relaxation time constant. These results suggest that the binding energy of nucleotide at NBD1 contributes to the overall energetics of the open channel conformation.
Figures


References
-
- Ai, T., S.G. Bompadre, X. Wang, S. Hu, M. Li, and T.-C. Hwang. 2004. Capsaicin potentiates wild-type and mutant CFTR chloride channel currents. Mol. Pharmacol. 65:1415–1426. - PubMed
-
- Aleksandrov, A.A., and J.R. Riordan. 1998. Regulation of CFTR ion channel gating by Mg-ATP. FEBS Lett. 431:97–101. - PubMed
