

Feline calicivirus VP2 is essential for the production of infectious virions
- PMID: 15767403
- PMCID: PMC1061574
- DOI: 10.1128/JVI.79.7.4012-4024.2005
Feline calicivirus VP2 is essential for the production of infectious virions
Abstract
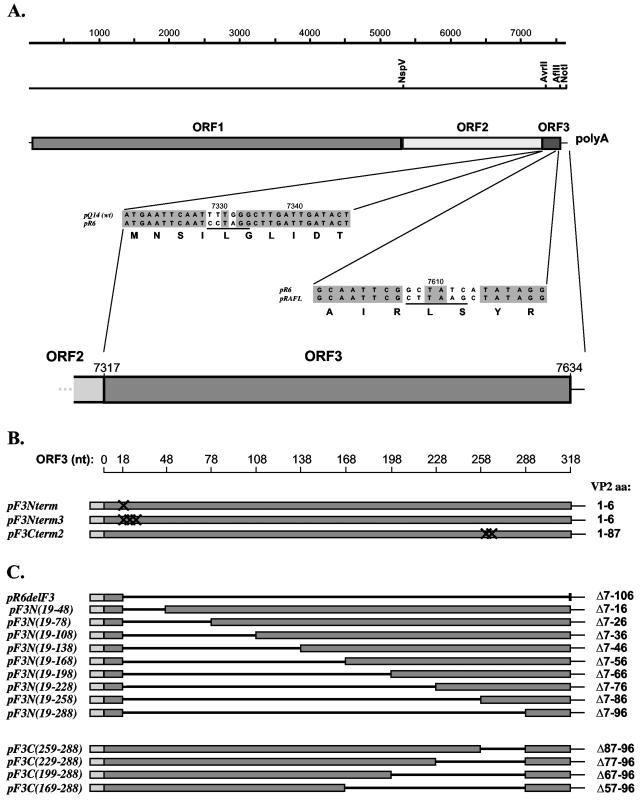
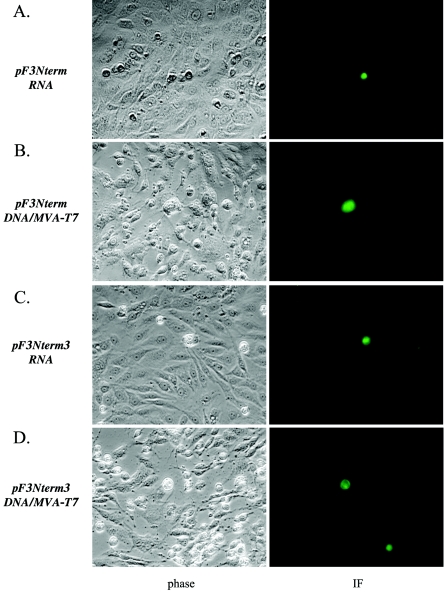
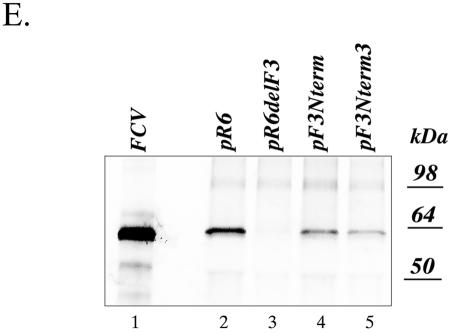
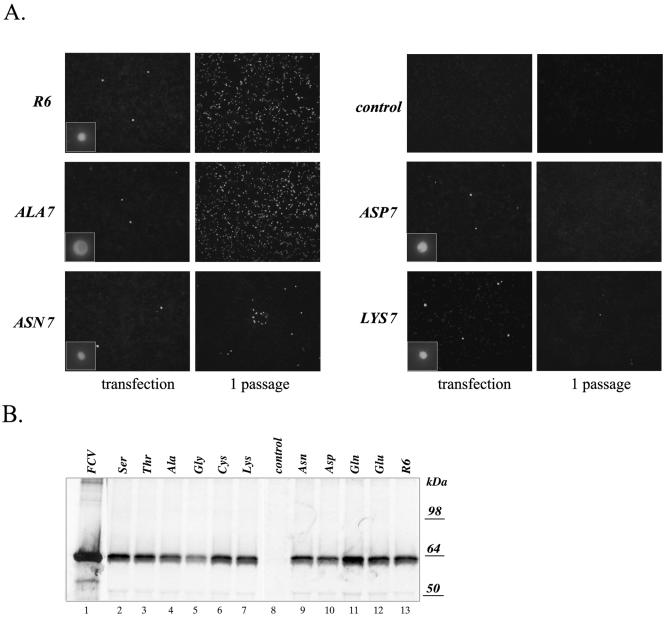
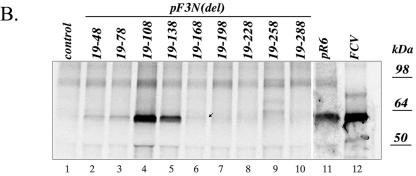
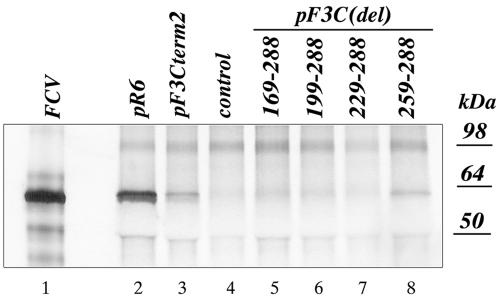
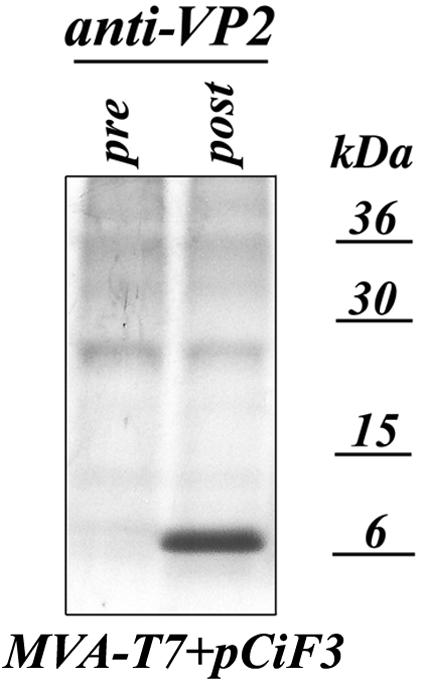
The third open reading frame (ORF3) located at the 3' end of the genomic RNA of feline calicivirus (FCV) encodes a small (12.2-kDa) minor structural protein of 106 amino acids designated VP2. Point mutations and deletions were introduced into an infectious FCV cDNA clone in order to evaluate the functional importance of ORF3 and its encoded protein, VP2. Deletion of the entire ORF3 sequence was lethal for the virus, and evidence was found for strong selective pressure to produce the VP2 protein. Extended deletions in the 5' end and small deletions in the 3' end of ORF3, as well as the introduction of stop codons into the ORF3 sequence, were tolerated by the viral replication machinery, but infectious virus could not be recovered. Infectious virus particles could be rescued from a full-length FCV cDNA clone encoding a nonfunctional VP2 when VP2 was provided in trans from a eukaryotic expression plasmid. Our data indicate that VP2, a protein apparently unique to the caliciviruses, is essential for productive replication that results in the synthesis and maturation of infectious virions and that the ORF3 nucleotide sequence itself overlaps a cis-acting RNA signal at the genomic 3' end.
Figures



References
-
- Burroughs, J. N., and F. Brown. 1978. Presence of a covalently linked protein on calicivirus RNA. J. Gen. Virol. 41:443-446. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
