

Evidence that CRABS CLAW and TOUSLED have conserved their roles in carpel development since the ancestor of the extant angiosperms
- PMID: 15767586
- PMCID: PMC555504
- DOI: 10.1073/pnas.0409577102
Evidence that CRABS CLAW and TOUSLED have conserved their roles in carpel development since the ancestor of the extant angiosperms
Abstract
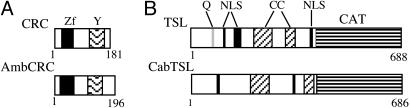
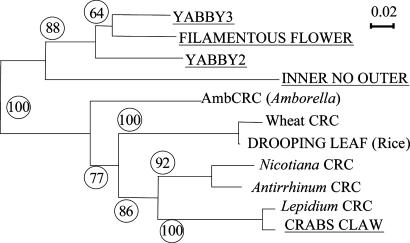
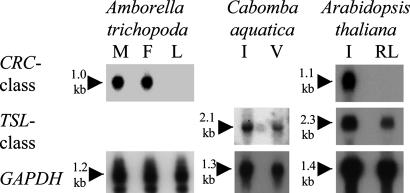
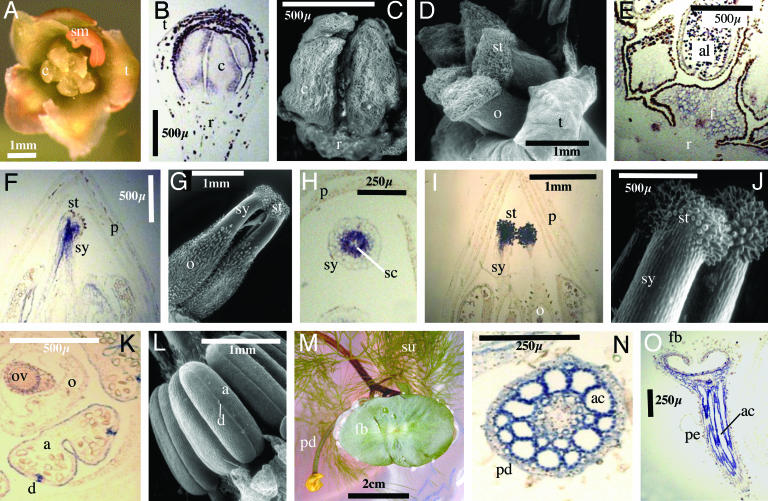
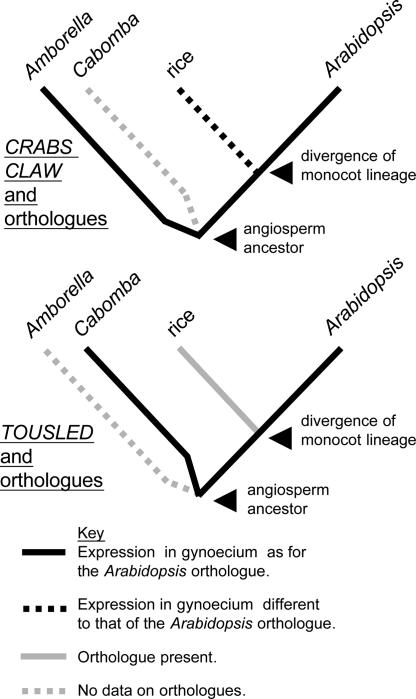
The carpel is the female reproductive organ specific to flowering plants. We aim to define the genes that controlled carpel development in the common ancestor of this group as a step toward determining the molecular events that were responsible for the evolution of the carpel. CRABS CLAW (CRC) and TOUSLED (TSL) control important aspects of carpel development in the model plant, Arabidopsis thaliana. The basal angiosperm species Amborella trichopoda and Cabomba aquatica very likely represent the two most early diverging groups of flowering plants. We have identified putative orthologues of CRC and TSL from A. trichopoda and C. aquatica, respectively. We demonstrate the expression patterns of these genes in carpels to be very highly conserved, both spatially and temporally, with those of their Arabidopsis orthologues. We argue that CRC and TSL in Arabidopsis are likely to have conserved their respective roles in carpel development since the common ancestor of the living flowering plants. We conclude that a divergent role shown for the CRC orthologue in rice, DROOPING LEAF, most probably arose specifically in the monocot lineage. We show that, in addition to its expression in carpels, the TSL orthologue of C. aquatica is expressed in tissues that contribute to buoyancy and argue that its role in these tissues may have arisen later than its role in carpel development.
Figures
Similar articles
-
Ancestral expression patterns and evolutionary diversification of YABBY genes in angiosperms.Plant J. 2011 Jul;67(1):26-36. doi: 10.1111/j.1365-313X.2011.04570.x. Epub 2011 Apr 26. Plant J. 2011. PMID: 21435049
-
Functional conservation between CRABS CLAW orthologues from widely diverged angiosperms.Ann Bot. 2007 Sep;100(3):651-7. doi: 10.1093/aob/mcm136. Epub 2007 Jul 23. Ann Bot. 2007. PMID: 17650514 Free PMC article.
-
The CRC orthologue from Pisum sativum shows conserved functions in carpel morphogenesis and vascular development.Ann Bot. 2014 Nov;114(7):1535-44. doi: 10.1093/aob/mcu129. Epub 2014 Jul 2. Ann Bot. 2014. PMID: 24989787 Free PMC article.
-
An evolutionary perspective on the regulation of carpel development.J Exp Bot. 2006;57(10):2143-52. doi: 10.1093/jxb/erj188. Epub 2006 May 23. J Exp Bot. 2006. PMID: 16720607 Review.
-
The formation and function of the female reproductive tract in flowering plants.Curr Biol. 2008 Oct 28;18(20):R972-8. doi: 10.1016/j.cub.2008.08.010. Curr Biol. 2008. PMID: 18957258 Review.
Cited by
-
The YABBY gene TONGARI-BOUSHI1 is involved in lateral organ development and maintenance of meristem organization in the rice spikelet.Plant Cell. 2012 Jan;24(1):80-95. doi: 10.1105/tpc.111.094797. Epub 2012 Jan 27. Plant Cell. 2012. PMID: 22286138 Free PMC article.
-
A light-regulated genetic module was recruited to carpel development in Arabidopsis following a structural change to SPATULA.Plant Cell. 2012 Jul;24(7):2812-25. doi: 10.1105/tpc.112.097915. Epub 2012 Jul 31. Plant Cell. 2012. PMID: 22851763 Free PMC article.
-
A physical map for the Amborella trichopoda genome sheds light on the evolution of angiosperm genome structure.Genome Biol. 2011;12(5):R48. doi: 10.1186/gb-2011-12-5-r48. Epub 2011 May 27. Genome Biol. 2011. PMID: 21619600 Free PMC article.
-
Flower development.Arabidopsis Book. 2010;8:e0127. doi: 10.1199/tab.0127. Epub 2010 Mar 23. Arabidopsis Book. 2010. PMID: 22303253 Free PMC article.
-
Morphological and molecular mechanisms of floral nectary development in Chinese Jujube.BMC Plant Biol. 2024 Nov 4;24(1):1041. doi: 10.1186/s12870-024-05760-8. BMC Plant Biol. 2024. PMID: 39497044 Free PMC article.
References
-
- Kuzoff, R. K. & Gasser, C. S. (2000) Trends Plant Sci. 5, 330–336. - PubMed
-
- Qiu, Y. L., Lee, J., Whitlock, B. A., Bernasconi-Quadroni, F. & Dombrovska, O. (2001) Mol. Biol. Evol. 18, 1745–1753. - PubMed
-
- Goremykin, V. V., Hirsch-Ernst, K. I., Wolfl, S. & Hellwig, F. H. (2003) Mol. Biol. Evol. 20, 1499–1505. - PubMed
-
- Goremykin, V. V., Hirsch-Ernst, K. I., Wolfl, S. & Hellwig, F. H. (2004) Mol. Biol. Evol. 21, 1445–1454. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
