

Kinase activation through dimerization by human SH2-B
- PMID: 15767667
- PMCID: PMC1061652
- DOI: 10.1128/MCB.25.7.2607-2621.2005
Kinase activation through dimerization by human SH2-B
Abstract
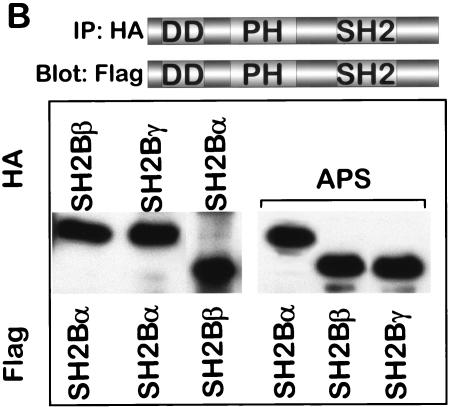
The isoforms of SH2-B, APS, and Lnk form a family of signaling proteins that have been described as activators, mediators, or inhibitors of cytokine and growth factor signaling. We now show that the three alternatively spliced isoforms of human SH2-B readily homodimerize in yeast two-hybrid and cellular transfections assays, and this is mediated specifically by a unique domain in its amino terminus. Consistent with previous reports, we further show that the SH2 domains of SH2-B and APS bind JAK2 at Tyr813. These findings suggested a model in which two molecules of SH2-B or APS homodimerize with their SH2 domains bound to two JAK2 molecules, creating heterotetrameric JAK2-(SH2-B)2-JAK2 or JAK2-(APS)2-JAK2 complexes. We further show that APS and SH2-B isoforms heterodimerize. At lower levels of SH2-B or APS expression, dimerization approximates two JAK2 molecules to induce transactivation. At higher relative concentrations of SH2-B or APS, kinase activation is blocked. SH2-B or APS homodimerization and SH2-B/APS heterodimerization thus provide direct mechanisms for activating and inhibiting JAK2 and other kinases from the inside of the cell and for potentiating or attenuating cytokine and growth factor receptor signaling when ligands are present.
Figures












Similar articles
-
Binding of SH2-B family members within a potential negative regulatory region maintains JAK2 in an active state.Mol Cell Biol. 2006 Sep;26(17):6381-94. doi: 10.1128/MCB.00570-06. Mol Cell Biol. 2006. PMID: 16914724 Free PMC article.
-
Interaction of fibroblast growth factor receptor 3 and the adapter protein SH2-B. A role in STAT5 activation.J Biol Chem. 2002 May 3;277(18):15962-70. doi: 10.1074/jbc.M102777200. Epub 2002 Feb 4. J Biol Chem. 2002. PMID: 11827956
-
Identification of SH2-bbeta as a potent cytoplasmic activator of the tyrosine kinase Janus kinase 2.Proc Natl Acad Sci U S A. 1999 Jun 22;96(13):7172-7. doi: 10.1073/pnas.96.13.7172. Proc Natl Acad Sci U S A. 1999. PMID: 10377387 Free PMC article.
-
SH2-B and SIRP: JAK2 binding proteins that modulate the actions of growth hormone.Recent Prog Horm Res. 2000;55:293-311. Recent Prog Horm Res. 2000. PMID: 11036942 Review.
-
Functional effects of APS and SH2-B on insulin receptor signalling.Biochem Soc Trans. 2001 Aug;29(Pt 4):529-34. doi: 10.1042/bst0290529. Biochem Soc Trans. 2001. PMID: 11498022 Review.
Cited by
-
The SH2B1 adaptor protein associates with a proximal region of the erythropoietin receptor.J Biol Chem. 2012 Jul 27;287(31):26223-34. doi: 10.1074/jbc.M112.382721. Epub 2012 Jun 5. J Biol Chem. 2012. PMID: 22669948 Free PMC article.
-
Binding of SH2-B family members within a potential negative regulatory region maintains JAK2 in an active state.Mol Cell Biol. 2006 Sep;26(17):6381-94. doi: 10.1128/MCB.00570-06. Mol Cell Biol. 2006. PMID: 16914724 Free PMC article.
-
Phosphorylation controls a dual-function polybasic nuclear localization sequence in the adapter protein SH2B1β to regulate its cellular function and distribution.J Cell Sci. 2011 May 1;124(Pt 9):1542-52. doi: 10.1242/jcs.078949. Epub 2011 Apr 12. J Cell Sci. 2011. PMID: 21486950 Free PMC article.
-
Adapter protein SH2B1beta cross-links actin filaments and regulates actin cytoskeleton.Mol Endocrinol. 2009 Jul;23(7):1065-76. doi: 10.1210/me.2008-0428. Epub 2009 Apr 2. Mol Endocrinol. 2009. PMID: 19342444 Free PMC article.
-
The effect of maternal care on gene expression and DNA methylation in a subsocial bee.Nat Commun. 2018 Aug 27;9(1):3468. doi: 10.1038/s41467-018-05903-0. Nat Commun. 2018. PMID: 30150650 Free PMC article.
References
-
- Bansal, A., and L. M. Gierasch. 1991. The NPXY internalization signal of the LDL receptor adopts a reverse-turn conformation. Cell 67:1195-1201. - PubMed
-
- Briddon, S. J., S. K. Melford, M. Turner, V. Tybulewicz, and S. P. Watson. 1999. Collagen mediates changes in intracellular calcium in primary mouse megakaryocytes through Syk-dependent and -independent pathways. Blood 93:3847-3855. - PubMed
-
- Carroll, D. J., D. T. Albay, M. Terasaki, L. A. Jaffe, and K. R. Foltz. 1999. Identification of PLCγ-dependent and -independent events during fertilization of sea urchin eggs. Dev. Biol. 206:232-247. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous