

Crossbridge properties investigated by fast ramp stretching of activated frog muscle fibres
- PMID: 15774512
- PMCID: PMC1464507
- DOI: 10.1113/jphysiol.2005.085209
Crossbridge properties investigated by fast ramp stretching of activated frog muscle fibres
Abstract
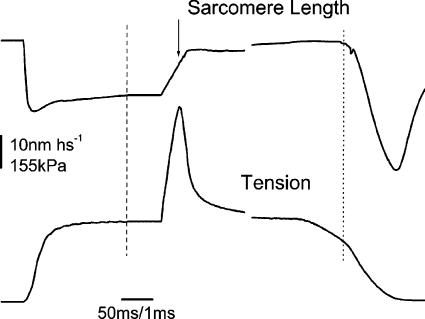
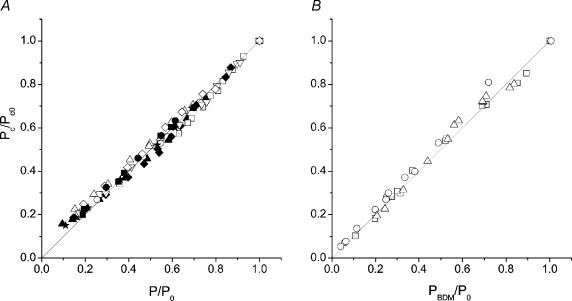
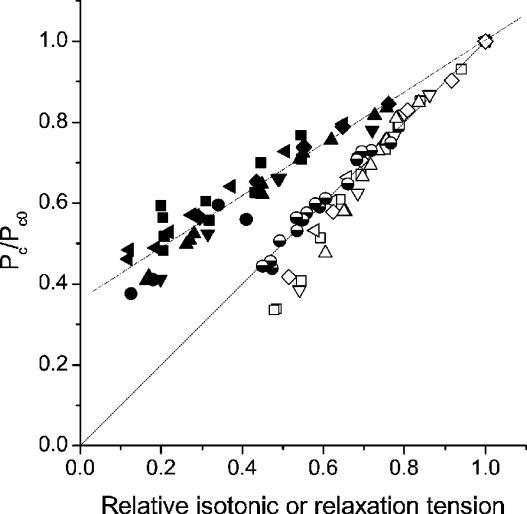
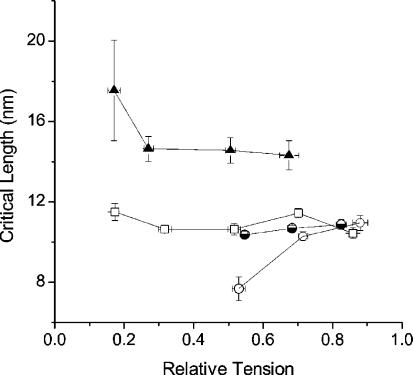
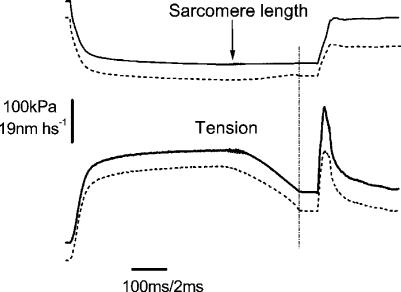
Very fast ramp stretches at 9.5-33 sarcomere lengths s(-1) (l0 s(-1)) stretching speed, 16-25 nm per half-sarcomere (nm hs(-1)) amplitude were applied to activated intact frog muscle fibres at tetanus plateau, during the tetanus rise, during the isometric phase of relaxation and during isotonic shortening. Stretches produced an almost linear tension increase above the isometric level up to a peak, and fell to a lower value in spite of continued stretching, indicating that the fibre became suddenly very compliant. This suggests that peak tension (critical tension, P(c)) represents the tension at which crossbridges are forcibly detached by the stretch. The ratio of P(c) to the isometric tension at tetanus plateau (P0) was 2.37 +/- 0.12 (S.E.M.). This ratio did not change significantly at lower tension (P) during the tetanus rise but decreased with time during the relaxation and increased with speed during isotonic shortening. At tetanus plateau P(c) occurred when sarcomere elongation attained a critical length (L(c)) of 10.98 +/- 0.13 nm hs(-1), independently of the stretching speed. L(c) remained constant during the tetanus rise but decreased on the relaxation and increased during isotonic shortening. Length-clamp experiments on the relaxation showed that the lower values of P(c)/P ratio and L(c), were both due to the slow sarcomere stretching occurring during this phase. Our data show that P(c) can be used as a measure of crossbridge number, while L(c) is a measure of crossbridge mean extension. Accordingly, for a given tension, crossbridges on the isometric relaxation are fewer than during the rise, develop a greater individual force and have a greater mean extension, while during isotonic shortening crossbridges are in a greater number but develop a smaller individual force and have a smaller extension.
Figures
Similar articles
-
Effects of solution tonicity on crossbridge properties and myosin lever arm disposition in intact frog muscle fibres.J Physiol. 2007 Jan 1;578(Pt 1):337-46. doi: 10.1113/jphysiol.2006.117770. Epub 2006 Oct 5. J Physiol. 2007. PMID: 17023505 Free PMC article.
-
Crossbridge properties during force enhancement by slow stretching in single intact frog muscle fibres.J Physiol. 2007 Dec 1;585(Pt 2):607-15. doi: 10.1113/jphysiol.2007.141440. Epub 2007 Oct 11. J Physiol. 2007. PMID: 17932153 Free PMC article.
-
Comparison of the tension responses to ramp shortening and lengthening in intact mammalian muscle fibres: crossbridge and non-crossbridge contributions.J Muscle Res Cell Motil. 2007;28(2-3):123-39. doi: 10.1007/s10974-007-9110-0. Epub 2007 Jul 4. J Muscle Res Cell Motil. 2007. PMID: 17610136
-
Crossbridge and non-crossbridge contributions to force in shortening and lengthening muscle.Adv Exp Med Biol. 2010;682:207-21. doi: 10.1007/978-1-4419-6366-6_12. Adv Exp Med Biol. 2010. PMID: 20824528 Review.
-
Non-crossbridge stiffness in active muscle fibres.J Exp Biol. 2016 Jan;219(Pt 2):153-60. doi: 10.1242/jeb.124370. J Exp Biol. 2016. PMID: 26792325 Review.
Cited by
-
Mechanism of force enhancement during stretching of skeletal muscle fibres investigated by high time-resolved stiffness measurements.J Muscle Res Cell Motil. 2013 Feb;34(1):71-81. doi: 10.1007/s10974-012-9335-4. Epub 2013 Jan 8. J Muscle Res Cell Motil. 2013. PMID: 23296372
-
The effects of Ca2+ and MgADP on force development during and after muscle length changes.PLoS One. 2013 Jul 16;8(7):e68866. doi: 10.1371/journal.pone.0068866. Print 2013. PLoS One. 2013. PMID: 23874795 Free PMC article.
-
Mechanism of force enhancement during and after lengthening of active muscle: a temperature dependence study.J Muscle Res Cell Motil. 2012 Oct;33(5):313-25. doi: 10.1007/s10974-012-9307-8. Epub 2012 Jun 16. J Muscle Res Cell Motil. 2012. PMID: 22706970
-
Sarcomere dynamics during muscular contraction and their implications to muscle function.J Muscle Res Cell Motil. 2007;28(1):89-104. doi: 10.1007/s10974-007-9107-8. Epub 2007 May 26. J Muscle Res Cell Motil. 2007. PMID: 17530424 Review.
-
Nonlinear Actomyosin Elasticity in Muscle?Biophys J. 2019 Jan 22;116(2):330-346. doi: 10.1016/j.bpj.2018.12.004. Epub 2018 Dec 13. Biophys J. 2019. PMID: 30606448 Free PMC article.
References
-
- Bagni MA, Cecchi G, Colombini B, Colomo F. Sarcomere tension-stiffness relation during the tetanus rise in single frog muscle fibres. J Muscle Res Cell Motil. 1999;20:469–476. - PubMed
-
- Bagni MA, Cecchi G, Colomo F, Garzella P. Effects of 2,3-butanedione monoxime on the crossbridge kinetics in frog single muscle fibres. J Muscle Res Cell Motil. 1992;13:516–522. - PubMed
-
- Bagni MA, Cecchi G, Colomo F, Tesi C. The mechanical characteristics of the contractile machinery at different levels of activation in intact single muscle fibres of the frog. Adv Exp Med Biol. 1988;226:473–487. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous