

Histone methyltransferases G9a and GLP form heteromeric complexes and are both crucial for methylation of euchromatin at H3-K9
- PMID: 15774718
- PMCID: PMC1074319
- DOI: 10.1101/gad.1284005
Histone methyltransferases G9a and GLP form heteromeric complexes and are both crucial for methylation of euchromatin at H3-K9
Abstract
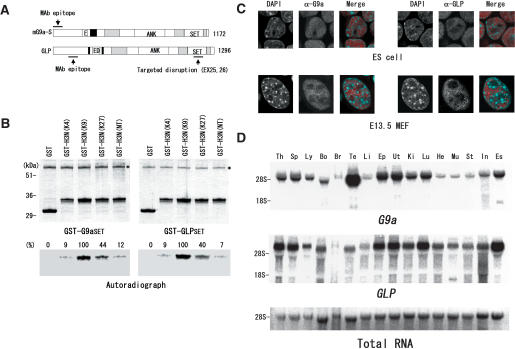
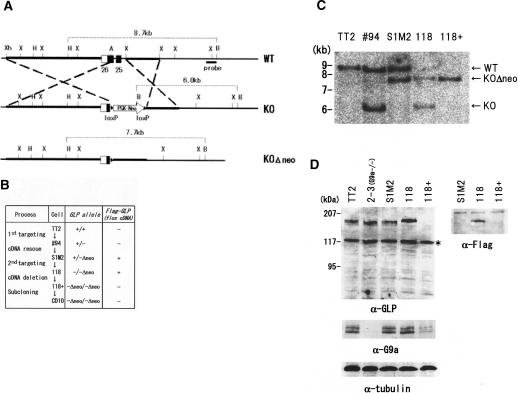
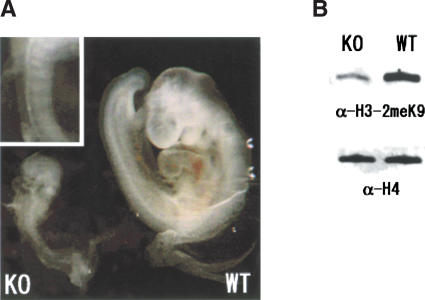
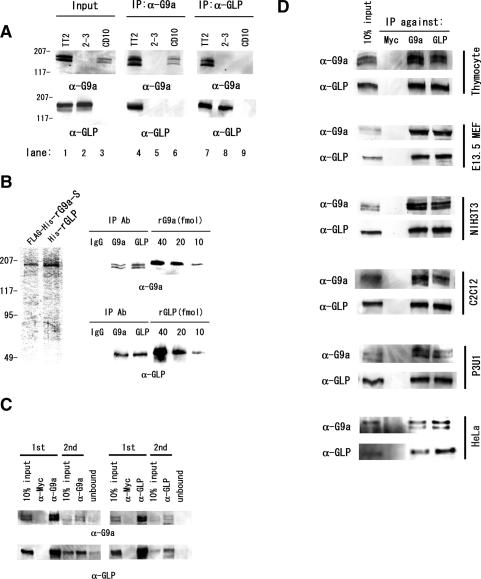
Histone H3 Lys 9 (H3-K9) methylation is a crucial epigenetic mark for transcriptional silencing. G9a is the major mammalian H3-K9 methyltransferase that targets euchromatic regions and is essential for murine embryogenesis. There is a single G9a-related methyltransferase in mammals, called GLP/Eu-HMTase1. Here we show that GLP is also important for H3-K9 methylation of mouse euchromatin. GLP-deficiency led to embryonic lethality, a severe reduction of H3-K9 mono- and dimethylation, the induction of Mage-a gene expression, and HP1 relocalization in embryonic stem cells, all of which were phenotypes of G9a-deficiency. Furthermore, we show that G9a and GLP formed a stoichiometric heteromeric complex in a wide variety of cell types. Biochemical analyses revealed that formation of the G9a/GLP complex was dependent on their enzymatic SET domains. Taken together, our new findings revealed that G9a and GLP cooperatively exert H3-K9 methyltransferase function in vivo, likely through the formation of higher-order heteromeric complexes.
Figures


References
-
- Aagaard L., Laible, G., Selenko, P., Schmid, M., Dorn, R., Schotta, G., Kuhfittig, S., Wolf, A., Lebersorger, A., Singh, P.B., et al. 1999. Functional mammalian homologs of the Drosophila PEV-modifier Su(var)3-9 encode centromere-associated proteins which complex with the heterochromatin component M31. EMBO J. 18: 1923–1938. - PMC - PubMed
-
- Bannister A.J., Zegerman, P., Partridge, J.F., Miska, E.A., Thomas, J.O., Allshire, R.C., and Kouzarides, T. 2001. Selective recognition of methylated lysine 9 on histone H3 by the HP1 chromo domain. Nature 410: 120–124. - PubMed
-
- Cao R. and Zhang, Y. 2004a. The functions of E(Z)/EZH2-mediated methylation of lysine 27 in histone H3. Curr. Opin. Genet. Dev. 14: 155–164. - PubMed
-
- ____. 2004b. SUZ12 is required for both the histone methyltransferase activity and the silencing function of the EED–EZH2 complex. Mol. Cell 15: 57–67. - PubMed
-
- Cheutin T., McNairn, A.J., Jenuwein, T., Gilbert, D.M., Singh, P.B., and Misteli, T. 2003. Maintenance of stable heterochromatin domains by dynamic HP1 binding. Science 299: 721–725. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials