

Analysis of the hypervariable region of the Salmonella enterica genome associated with tRNA(leuX)
- PMID: 15774890
- PMCID: PMC1065210
- DOI: 10.1128/JB.187.7.2469-2482.2005
Analysis of the hypervariable region of the Salmonella enterica genome associated with tRNA(leuX)
Abstract
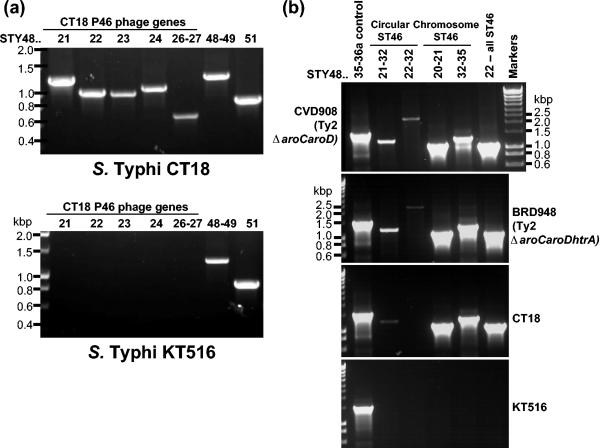
The divergence of Salmonella enterica and Escherichia coli is estimated to have occurred approximately 140 million years ago. Despite this evolutionary distance, the genomes of these two species still share extensive synteny and homology. However, there are significant differences between the two species in terms of genes putatively acquired via various horizontal transfer events. Here we report on the composition and distribution across the Salmonella genus of a chromosomal region designated SPI-10 in Salmonella enterica serovar Typhi and located adjacent to tRNA(leuX). We find that across the Salmonella genus the tRNA(leuX) region is a hypervariable hot spot for horizontal gene transfer; different isolates from the same S. enterica serovar can exhibit significant variation in this region. Many P4 phage, plasmid, and transposable element-associated genes are found adjacent to tRNA(leuX) in both Salmonella and E. coli, suggesting that these mobile genetic elements have played a major role in driving the variability of this region.
Figures





References
-
- Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990. Basic local alignment search tool. J. Mol. Biol. 215:403-410. - PubMed
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, and J. A. Smith. 1995. Preparation of genomic DNA from bacteria, p. 2.11-2.22. In K. Struhl (ed.), Short protocols in molecular biology, vol. 3. John Wiley and Sons, Inc., New York, N.Y.
-
- Baumler, A. J., A. J. Gilde, R. M. Tsolis, A. W. van der Velden, B. M. Ahmer, and F. Heffron. 1997. Contribution of horizontal gene transfer and deletion events to development of distinctive patterns of fimbrial operons during evolution of Salmonella serotypes. J. Bacteriol. 179:317-322. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
