

LdpA: a component of the circadian clock senses redox state of the cell
- PMID: 15775978
- PMCID: PMC556408
- DOI: 10.1038/sj.emboj.7600606
LdpA: a component of the circadian clock senses redox state of the cell
Abstract
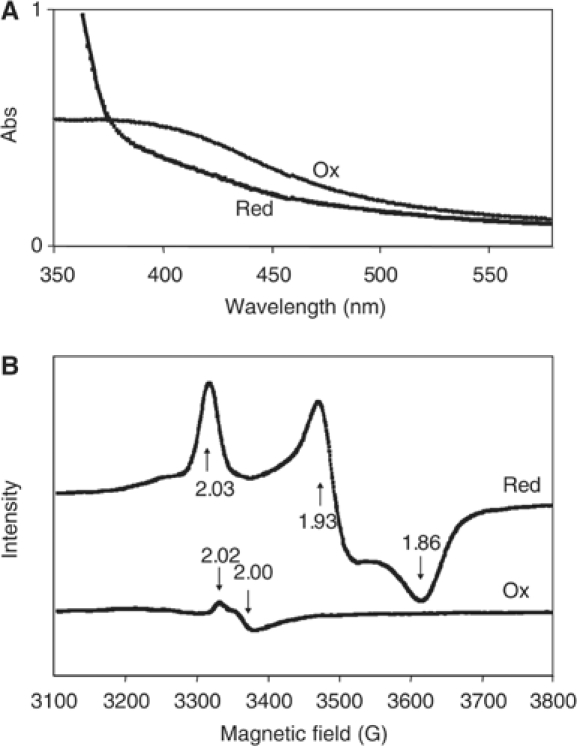
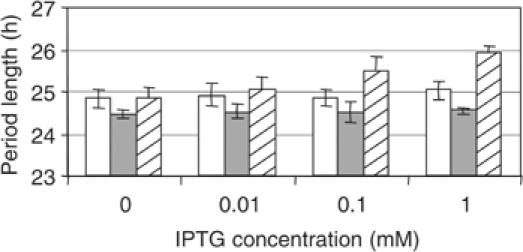
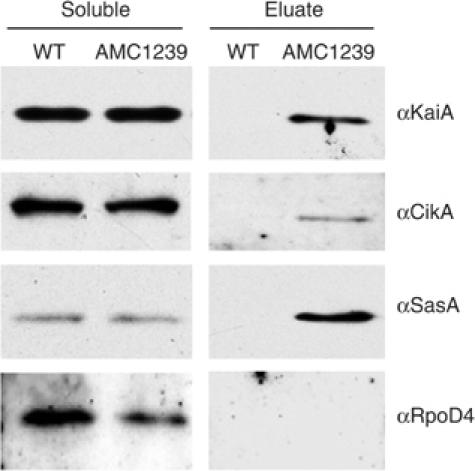
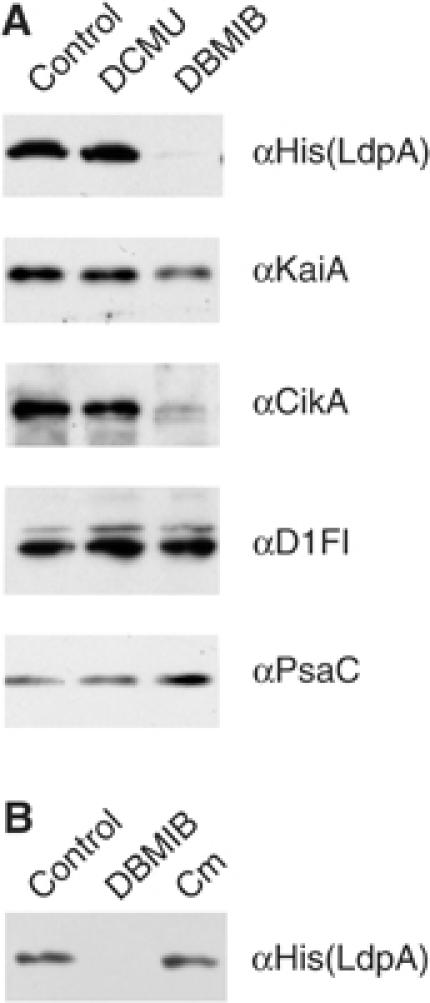
The endogenous 24-h (circadian) rhythms exhibited by the cyanobacterium Synechococcus elongatus PCC 7942 and other organisms are entrained by a variety of environmental factors. In cyanobacteria, the mechanism that transduces environmental input signals to the central oscillator of the clock is not known. An earlier study identified ldpA as a gene involved in light-dependent modulation of the circadian period, and a candidate member of a clock-entraining input pathway. Here, we report that the LdpA protein is sensitive to the redox state of the cell and exhibits electron paramagnetic resonance spectra consistent with the presence of two Fe4S4 clusters. Moreover, LdpA copurifies with proteins previously shown to be integral parts of the circadian mechanism. We also demonstrate that LdpA affects both the absolute level and light-dependent variation in abundance of CikA, a key input pathway component. The data suggest a novel input pathway to the circadian oscillator in which LdpA is a component of the clock protein complex that senses the redox state of a cell.
Figures
References
-
- Aschoff J (1981) Freerunning and entrained circadian rhythms. In Handbook of Behavioral Neurobiology: Biological Rhythms, Aschoff J (ed) pp 81–93. NY: Plenum Press
-
- Barnes D, Mayfield SP (2003) Redox control of posttranscriptional processes in the chloroplast. Antioxid Redox Signal 5: 89–94 - PubMed
-
- Beinert H, Kiley PJ (1999) Fe-S proteins in sensing and regulatory functions. Curr Opin Chem Biol 3: 152–157 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
