

Invasion of Lotus japonicus root hairless 1 by Mesorhizobium loti involves the nodulation factor-dependent induction of root hairs
- PMID: 15778455
- PMCID: PMC1088324
- DOI: 10.1104/pp.104.057513
Invasion of Lotus japonicus root hairless 1 by Mesorhizobium loti involves the nodulation factor-dependent induction of root hairs
Abstract
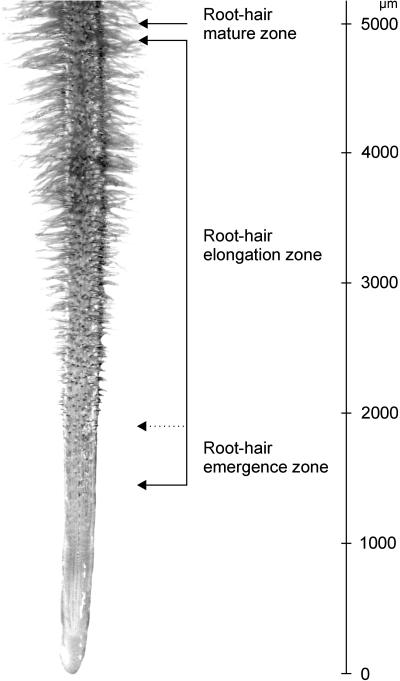
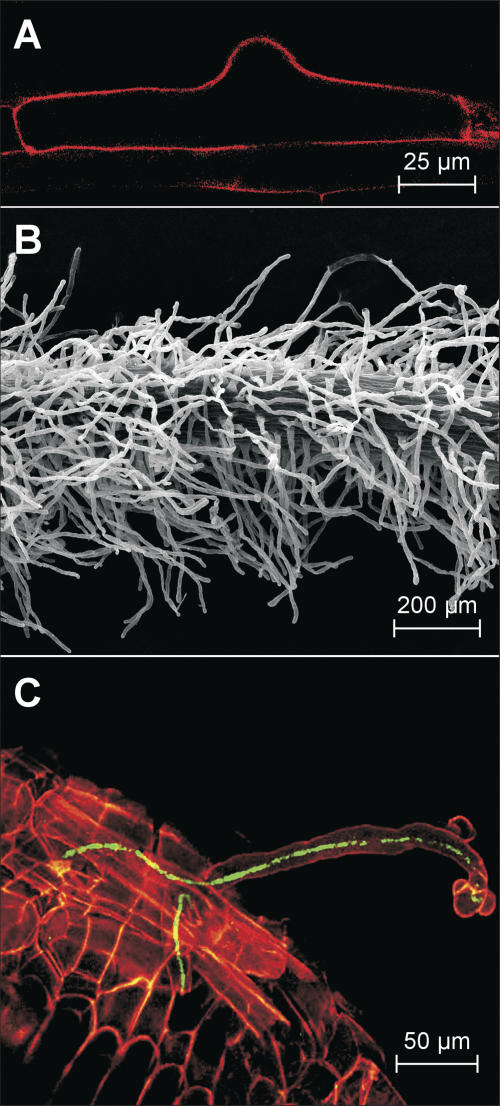
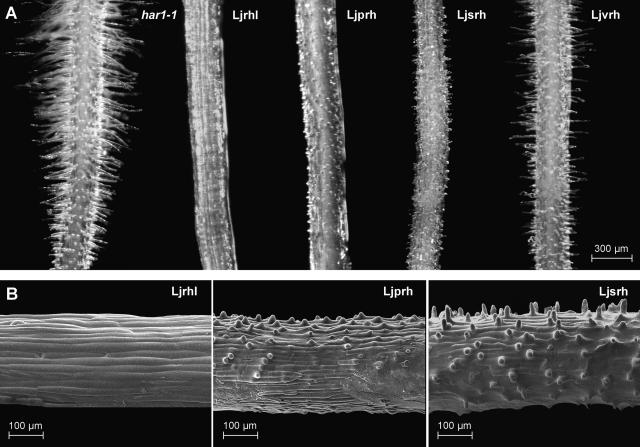
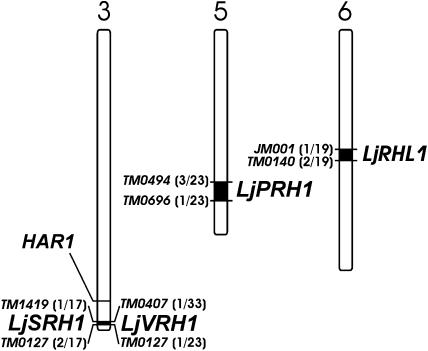
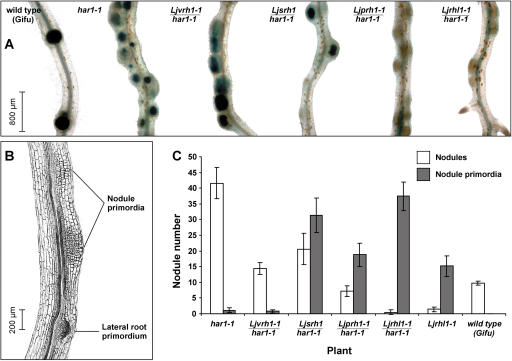
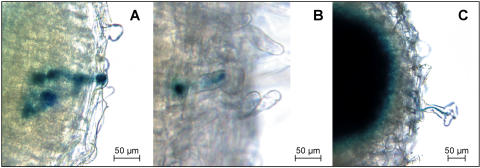
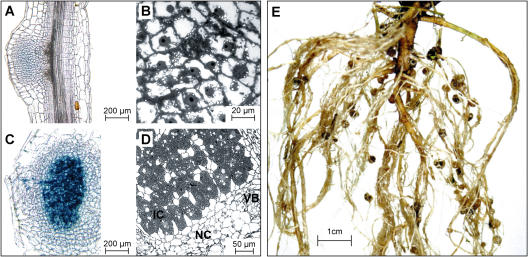
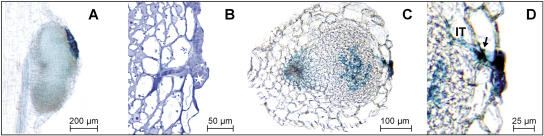
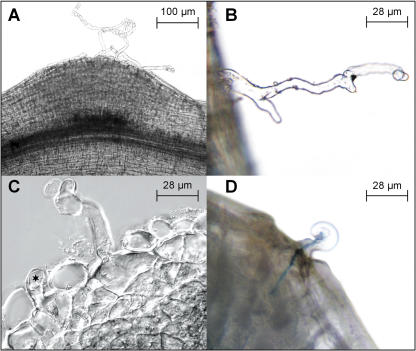
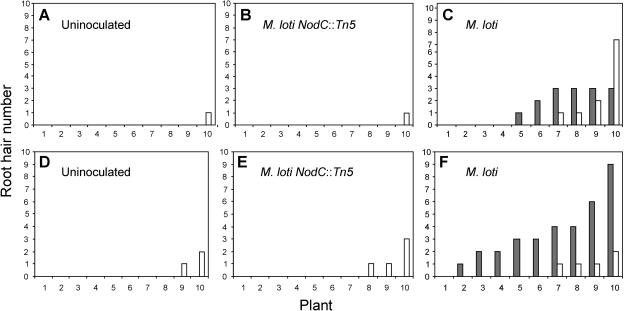
In many legumes, including Lotus japonicus and Medicago truncatula, susceptible root hairs are the primary sites for the initial signal perception and physical contact between the host plant and the compatible nitrogen-fixing bacteria that leads to the initiation of root invasion and nodule organogenesis. However, diverse mechanisms of nodulation have been described in a variety of legume species that do not rely on root hairs. To clarify the significance of root hairs during the L. japonicus-Mesorhizobium loti symbiosis, we have isolated and performed a detailed analysis of four independent L. japonicus root hair developmental mutants. We show that although important for the efficient colonization of roots, the presence of wild-type root hairs is not required for the initiation of nodule primordia (NP) organogenesis and the colonization of the nodule structures. In the genetic background of the L. japonicus root hairless 1 mutant, the nodulation factor-dependent formation of NP provides the structural basis for alternative modes of invasion by M. loti. Surprisingly, one mode of root colonization involves nodulation factor-dependent induction of NP-associated cortical root hairs and epidermal root hairs, which, in turn, support bacterial invasion. In addition, entry of M. loti through cracks at the cortical surface of the NP is described. These novel mechanisms of nodule colonization by M. loti explain the fully functional, albeit significantly delayed, nodulation phenotype of the L. japonicus ROOT HAIRLESS mutant.
Figures
Similar articles
-
Grafting between model legumes demonstrates roles for roots and shoots in determining nodule type and host/rhizobia specificity.J Exp Bot. 2005 Jun;56(416):1643-50. doi: 10.1093/jxb/eri160. Epub 2005 Apr 11. J Exp Bot. 2005. PMID: 15824071
-
New nodulation mutants responsible for infection thread development in Lotus japonicus.Mol Plant Microbe Interact. 2006 Jul;19(7):801-10. doi: 10.1094/MPMI-19-0801. Mol Plant Microbe Interact. 2006. PMID: 16838792
-
crinkle, a novel symbiotic mutant that affects the infection thread growth and alters the root hair, trichome, and seed development in Lotus japonicus.Plant Physiol. 2003 Mar;131(3):1054-63. doi: 10.1104/pp.102.017020. Plant Physiol. 2003. PMID: 12644658 Free PMC article.
-
Leguminous plants: inventors of root nodules to accommodate symbiotic bacteria.Int Rev Cell Mol Biol. 2015;316:111-58. doi: 10.1016/bs.ircmb.2015.01.004. Epub 2015 Feb 20. Int Rev Cell Mol Biol. 2015. PMID: 25805123 Review.
-
Sesbania rostrata: a case study of natural variation in legume nodulation.New Phytol. 2010 Apr;186(2):340-5. doi: 10.1111/j.1469-8137.2009.03124.x. Epub 2009 Dec 15. New Phytol. 2010. PMID: 20015069 Review.
Cited by
-
The RPG gene of Medicago truncatula controls Rhizobium-directed polar growth during infection.Proc Natl Acad Sci U S A. 2008 Jul 15;105(28):9817-22. doi: 10.1073/pnas.0710273105. Epub 2008 Jul 9. Proc Natl Acad Sci U S A. 2008. PMID: 18621693 Free PMC article.
-
Rearrangement of actin cytoskeleton mediates invasion of Lotus japonicus roots by Mesorhizobium loti.Plant Cell. 2009 Jan;21(1):267-84. doi: 10.1105/tpc.108.063693. Epub 2009 Jan 9. Plant Cell. 2009. PMID: 19136645 Free PMC article.
-
Root hairs are the most important root trait for rhizosheath formation of barley (Hordeum vulgare), maize (Zea mays) and Lotus japonicus (Gifu).Ann Bot. 2021 Jul 28;128(1):45-57. doi: 10.1093/aob/mcab029. Ann Bot. 2021. PMID: 33631013 Free PMC article.
-
Conservation of lotus and Arabidopsis basic helix-loop-helix proteins reveals new players in root hair development.Plant Physiol. 2009 Nov;151(3):1175-85. doi: 10.1104/pp.109.143867. Epub 2009 Aug 12. Plant Physiol. 2009. PMID: 19675148 Free PMC article.
-
The molecular network governing nodule organogenesis and infection in the model legume Lotus japonicus.Nat Commun. 2010 Apr 12;1(1):10. doi: 10.1038/ncomms1009. Nat Commun. 2010. PMID: 20975672 Free PMC article.
References
-
- Ardourel M, Demont N, Debelle F, Maillet F, de Billy F, Prome J-C, Denarie J, Truchet G (1994) Rhizobium meliloti lipooligosaccharide nodulation factors: different structural requirements for bacterial entry into target root hair cells and induction of plant symbiotic developmental responses. Plant Cell 6: 1357–1374 - PMC - PubMed
-
- Asamizu E, Kato T, Sato S, Nakamura Y, Kaneko T, Tabata S (2003) Structural analysis of a Lotus japonicus genome. IV. Sequence features and mapping of seventy-three TAC clones which cover the 7.5 Mb regions of the genome. DNA Res 10: 115–122 - PubMed
-
- Boogerd FC, van Rossum D (1997) Nodulation of groundnut by Bradyrhizobium: a simple infection process by crack entry. FEMS Microbiol Rev 21: 5–27
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
