

Tissue-specific localization of pea root infection by Nectria haematococca. Mechanisms and consequences
- PMID: 15778461
- PMCID: PMC1088327
- DOI: 10.1104/pp.104.056366
Tissue-specific localization of pea root infection by Nectria haematococca. Mechanisms and consequences
Abstract
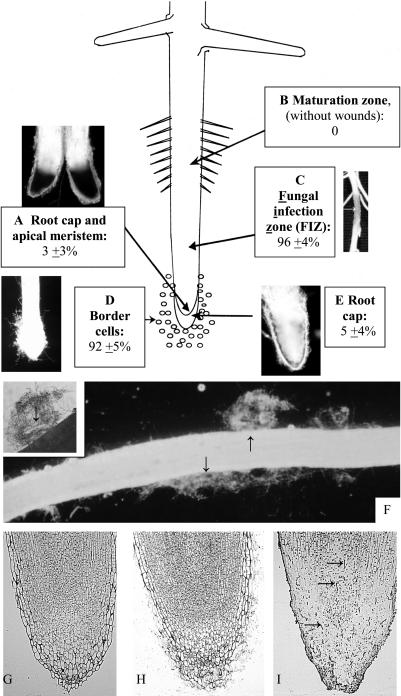
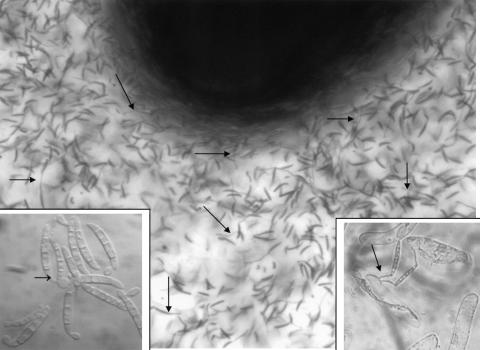
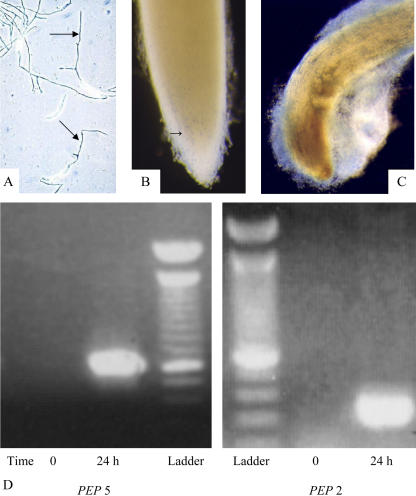
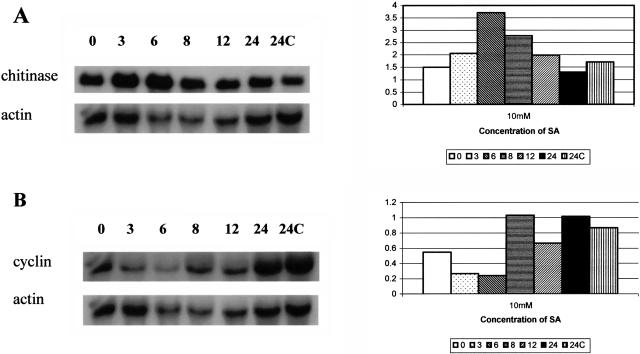
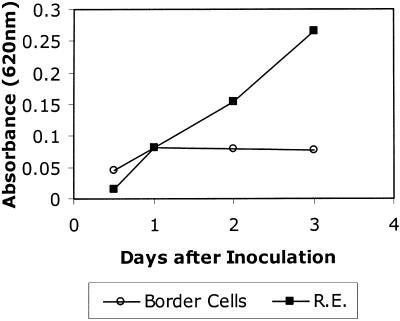
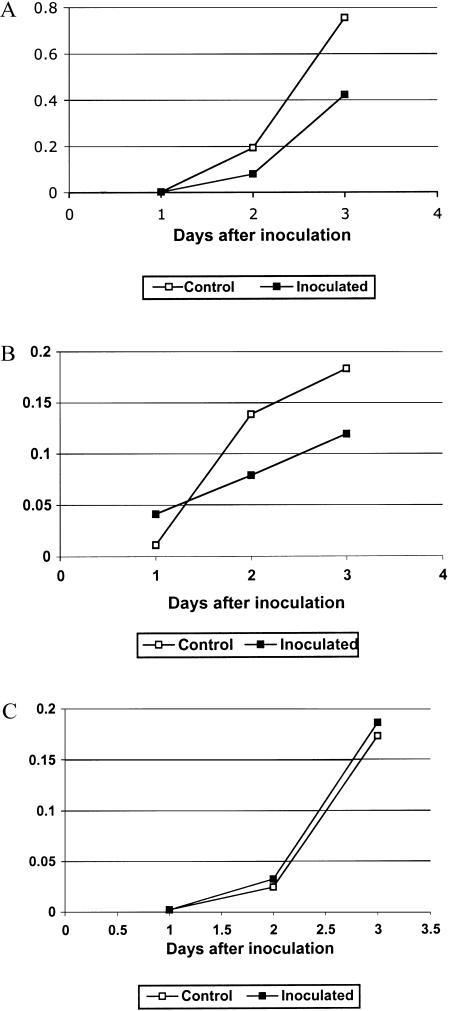
Root infection in susceptible host species is initiated predominantly in the zone of elongation, whereas the remainder of the root is resistant. Nectria haematococca infection of pea (Pisum sativum) was used as a model to explore possible mechanisms influencing the localization of root infection. The failure to infect the root tip was not due to a failure to induce spore germination at this site, suppression of pathogenicity genes in the fungus, or increased expression of plant defense genes. Instead, exudates from the root tip induce rapid spore germination by a pathway that is independent of nutrient-induced germination. Subsequently, a factor produced during fungal infection and death of border cells at the root apex appears to selectively suppress fungal growth and prevent sporulation. Host-specific mantle formation in response to border cells appears to represent a previously unrecognized form of host-parasite relationship common to diverse species. The dynamics of signal exchange leading to mantle development may play a key role in fostering plant health, by protecting root meristems from pathogenic invasion.
Figures
Similar articles
-
Introduction of plant and fungal genes into pea (Pisum sativum L.) hairy roots reduces their ability to produce pisatin and affects their response to a fungal pathogen.Mol Plant Microbe Interact. 2004 Jul;17(7):798-804. doi: 10.1094/MPMI.2004.17.7.798. Mol Plant Microbe Interact. 2004. PMID: 15242174
-
Tissue specific localization of root infection by fungal pathogens: role of root border cells.Mol Plant Microbe Interact. 2002 Nov;15(11):1128-36. doi: 10.1094/MPMI.2002.15.11.1128. Mol Plant Microbe Interact. 2002. PMID: 12423018
-
Extracellular DNA is required for root tip resistance to fungal infection.Plant Physiol. 2009 Oct;151(2):820-9. doi: 10.1104/pp.109.142067. Epub 2009 Aug 21. Plant Physiol. 2009. PMID: 19700564 Free PMC article.
-
Host recognition by pathogenic fungi through plant flavonoids.Adv Exp Med Biol. 2002;505:9-22. doi: 10.1007/978-1-4757-5235-9_2. Adv Exp Med Biol. 2002. PMID: 12083470 Review.
-
In vitro transcription and binding analysis of promoter regulation by a host-specific signal in a phytopathogenic fungus.Antonie Van Leeuwenhoek. 1994;65(3):183-9. doi: 10.1007/BF00871945. Antonie Van Leeuwenhoek. 1994. PMID: 7847884 Review.
Cited by
-
Root exudate of Solanum tuberosum is enriched in galactose-containing molecules and impacts the growth of Pectobacterium atrosepticum.Ann Bot. 2016 Oct 1;118(4):797-808. doi: 10.1093/aob/mcw128. Ann Bot. 2016. PMID: 27390353 Free PMC article.
-
The production and release of living root cap border cells is a function of root apical meristem type in dicotyledonous angiosperm plants.Ann Bot. 2006 May;97(5):917-23. doi: 10.1093/aob/mcj602. Epub 2006 Feb 17. Ann Bot. 2006. PMID: 16488922 Free PMC article.
-
Proteins among the polysaccharides: a new perspective on root cap slime.Plant Signal Behav. 2007 Sep;2(5):410-2. doi: 10.4161/psb.2.5.4344. Plant Signal Behav. 2007. PMID: 19704617 Free PMC article.
-
Plant Immune Mechanisms: From Reductionistic to Holistic Points of View.Mol Plant. 2020 Oct 5;13(10):1358-1378. doi: 10.1016/j.molp.2020.09.007. Epub 2020 Sep 8. Mol Plant. 2020. PMID: 32916334 Free PMC article. Review.
-
Root Border Cells and Mucilage Secretions of Soybean, Glycine Max (Merr) L.: Characterization and Role in Interactions with the Oomycete Phytophthora Parasitica.Cells. 2020 Sep 30;9(10):2215. doi: 10.3390/cells9102215. Cells. 2020. PMID: 33008016 Free PMC article.
References
-
- Arriola L, Niemira BA, Safir GR (1997) Border cells and arbuscular mycorrhizae in four Amaranthaceae species. Phytopathology 87: 1240–1242 - PubMed
-
- Atkinson TG, Neal JL, Larson RI (1975) Genetic control of the rhizosphere microflora of wheat. In GW Bruehl, ed, Biology and Control of Soil-borne Plant Pathogens. American Phytopathological Society, St. Paul, pp 116–122
-
- Ayers WA, Thornton RH (1968) Exudation of amino acids by intact and damaged roots of wheat and peas. Plant Soil 28: 193–207
-
- Bar-Yosef B (1996) Root excretions and their environmental effects: influence on availability of phosphorus. In Y Waisel, A Eshel, U Kafkafi, eds, Plant Roots: The Hidden Half. Marcel Dekker, New York, pp 581–606
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
