

Spike protein assembly into the coronavirion: exploring the limits of its sequence requirements
- PMID: 15780881
- PMCID: PMC7111810
- DOI: 10.1016/j.virol.2005.02.001
Spike protein assembly into the coronavirion: exploring the limits of its sequence requirements
Abstract
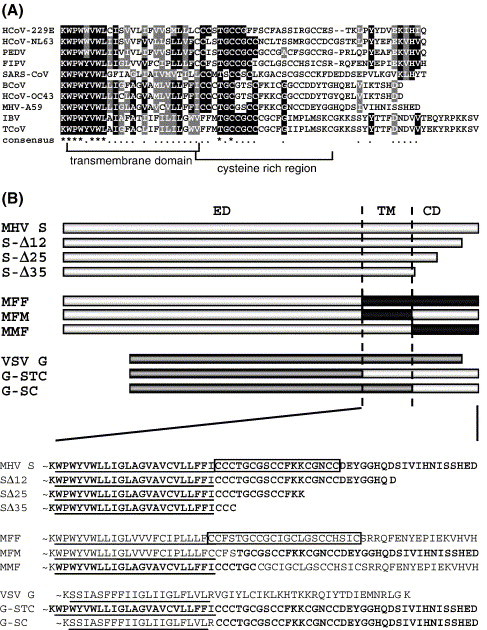
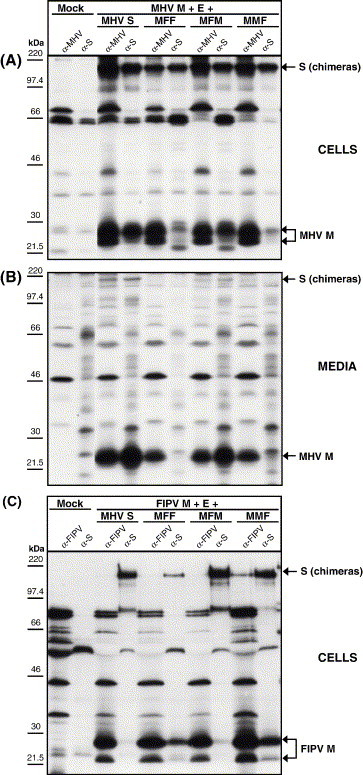
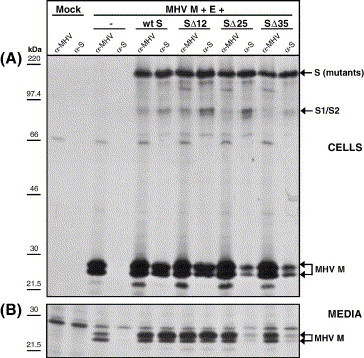
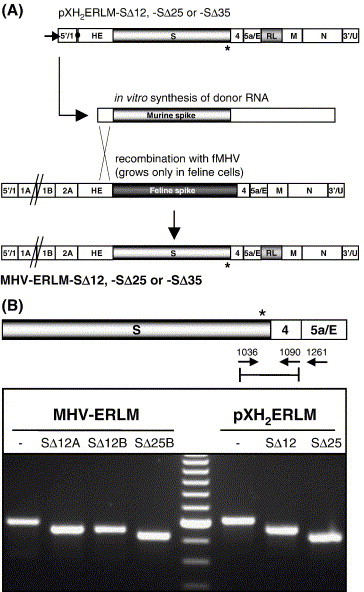
The coronavirus spike (S) protein, required for receptor binding and membrane fusion, is incorporated into the assembling virion by interactions with the viral membrane (M) protein. Earlier we showed that the ectodomain of the S protein is not involved in this process. Here we further defined the requirements of the S protein for virion incorporation. We show that the cytoplasmic domain, not the transmembrane domain, determines the association with the M protein and suffices to effect the incorporation into viral particles of chimeric spikes as well as of foreign viral glycoproteins. The essential sequence was mapped to the membrane-proximal region of the cytoplasmic domain, which is also known to be of critical importance for the fusion function of the S protein. Consistently, only short C-terminal truncations of the S protein were tolerated when introduced into the virus by targeted recombination. The important role of the about 38-residues cytoplasmic domain in the assembly of and membrane fusion by this approximately 1300 amino acids long protein is discussed.
Figures
References
-
- Ali A., Nayak D.P. Assembly of Sendai virus: M protein interacts with F and HN proteins and with the cytoplasmic tail and transmembrane domain of F protein. Virology. 2000;276(2):289–303. - PubMed
-
- Cavanagh D. The coronavirus surface glycoprotein. In: Siddell S.G., editor. The coronaviridae. Plenum Press; New York: 1995.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
