

Both RNase E and RNase III control the stability of sodB mRNA upon translational inhibition by the small regulatory RNA RyhB
- PMID: 15781494
- PMCID: PMC1069011
- DOI: 10.1093/nar/gki313
Both RNase E and RNase III control the stability of sodB mRNA upon translational inhibition by the small regulatory RNA RyhB
Abstract
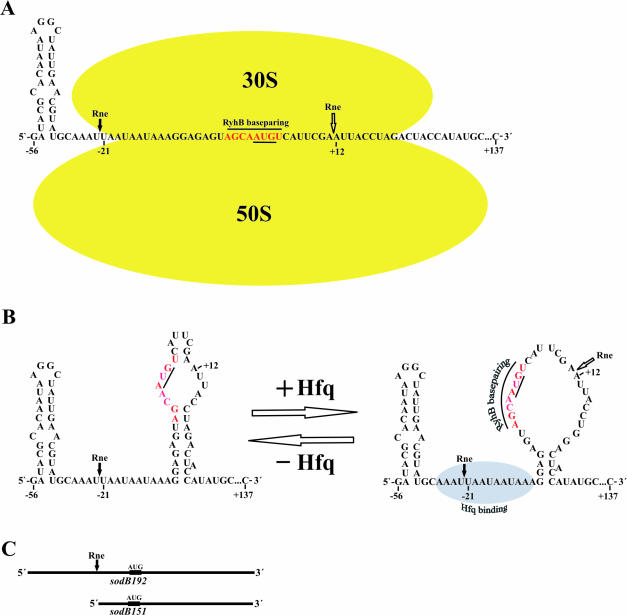
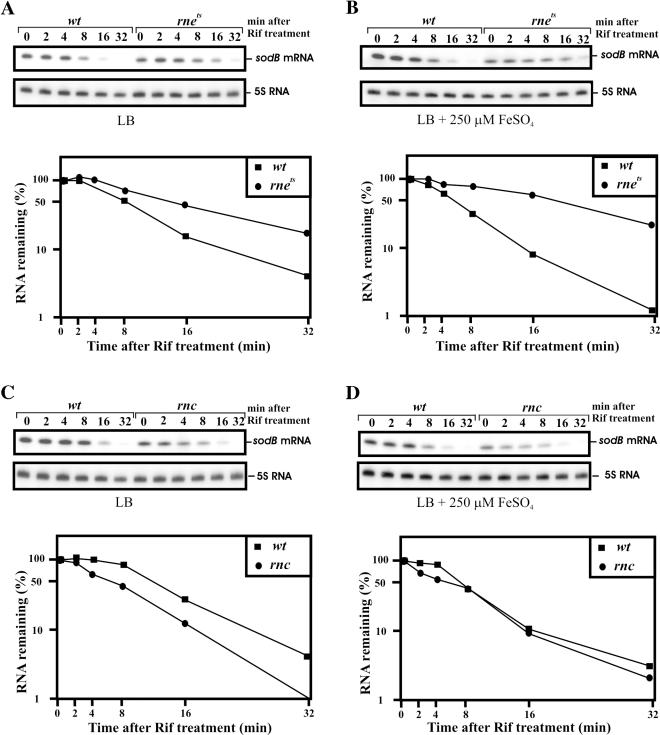
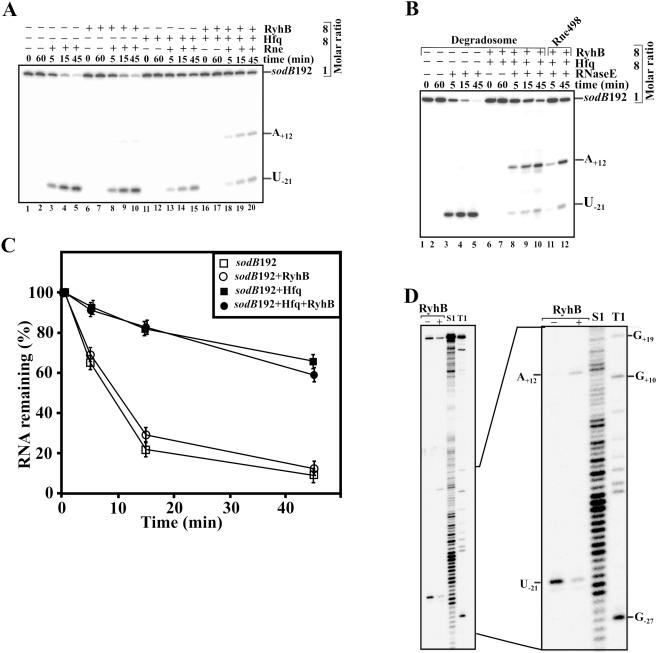
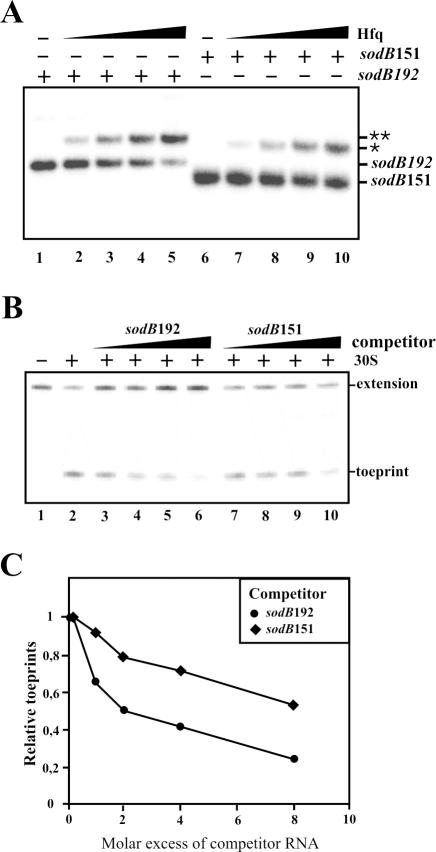
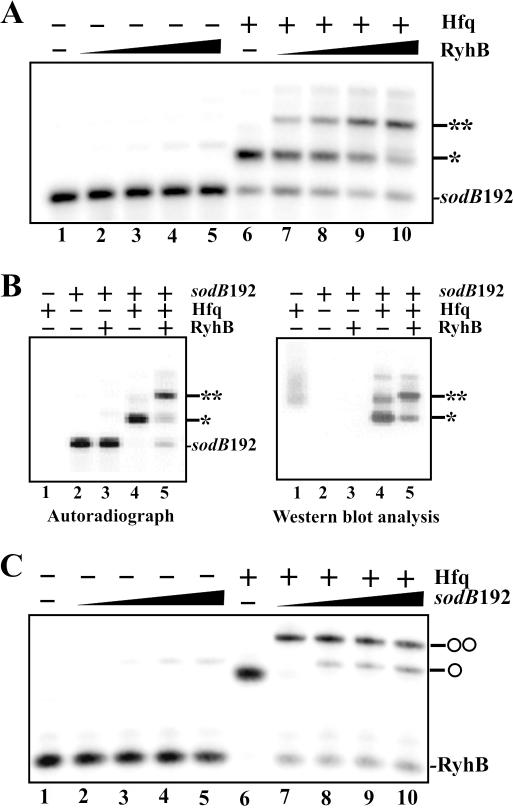
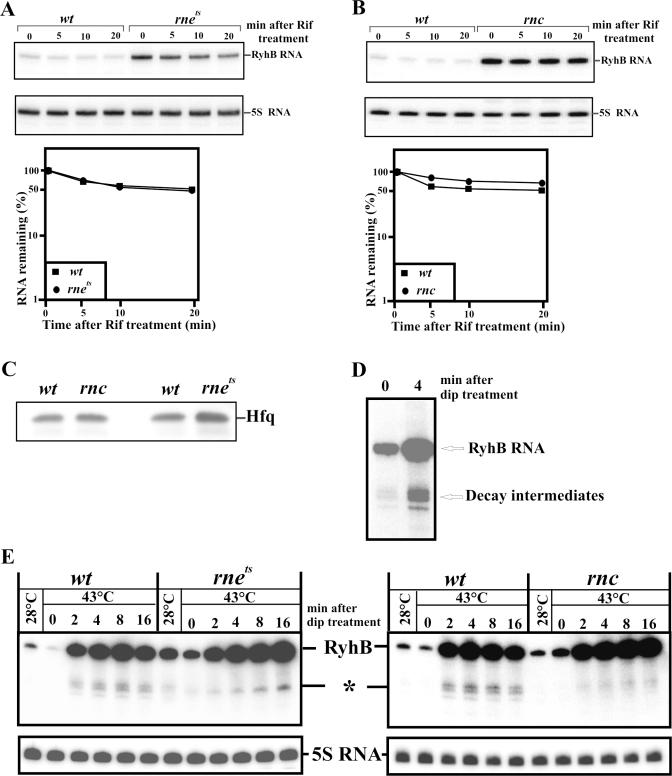
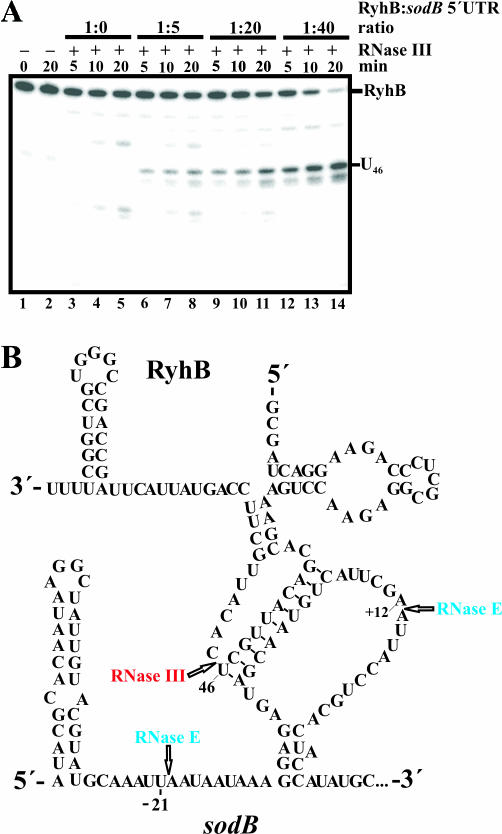
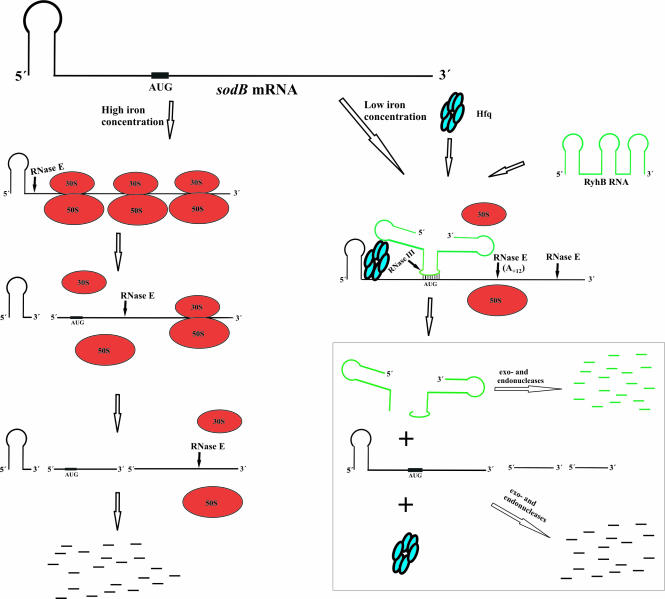
Previous work has demonstrated that iron-dependent variations in the steady-state concentration and translatability of sodB mRNA are modulated by the small regulatory RNA RyhB, the RNA chaperone Hfq and RNase E. In agreement with the proposed role of RNase E, we found that the decay of sodB mRNA is retarded upon inactivation of RNase E in vivo, and that the enzyme cleaves within the sodB 5'-untranslated region (5'-UTR) in vitro, thereby removing the 5' stem-loop structure that facilitates Hfq and ribosome binding. Moreover, RNase E cleavage can also occur at a cryptic site that becomes available upon sodB 5'-UTR/RyhB base pairing. We show that while playing an important role in facilitating the interaction of RyhB with sodB mRNA, Hfq is not tightly retained by the RyhB-sodB mRNA complex and can be released from it through interaction with other RNAs added in trans. Unlike turnover of sodB mRNA, RyhB decay in vivo is mainly dependent on RNase III, and its cleavage by RNase III in vitro is facilitated upon base pairing with the sodB 5'-UTR. These data are discussed in terms of a model, which accounts for the observed roles of RNase E and RNase III in sodB mRNA turnover.
Figures
References
-
- Le Derout J., Regnier P., Hajnsdorf E. Both temperature and medium composition regulate RNase E processing efficiency of the rpsO mRNA coding for ribosomal protein S15 of Escherichia coli. J. Mol. Biol. 2002;319:341–349. - PubMed
-
- Nilsson G., Belasco J.G., Cohen S.N., von Gabain A. Growth-rate dependent regulation of mRNA stability in Escherichia coli. Nature. 1984;312:75–77. - PubMed
-
- Coburn G.A., Mackie G.A. Degradation of mRNA in Escherichia coli: an old problem with some new twists. Prog. Nucleic Acid Res. Mol. Biol. 1999;62:55–108. - PubMed
-
- Lundberg U., Kaberdin V., von Gabain A. The mechanisms of mRNA degradation in bacteria and their implication for stabilization of heterologous transcripts. In: Demain A.L., Davies R.M., Cohen G., Hershberg C.L., Sherman D.H., Willson R.C., Wu J.-H.D., editors. Manual of Industrial Microbiology and Biotechnology. 2nd edn. Washington, DC: ASM Press; 1999. pp. 585–596.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
