

Roles of polymerization dynamics, opposed motors, and a tensile element in governing the length of Xenopus extract meiotic spindles
- PMID: 15788560
- PMCID: PMC1142448
- DOI: 10.1091/mbc.e05-02-0174
Roles of polymerization dynamics, opposed motors, and a tensile element in governing the length of Xenopus extract meiotic spindles
Abstract
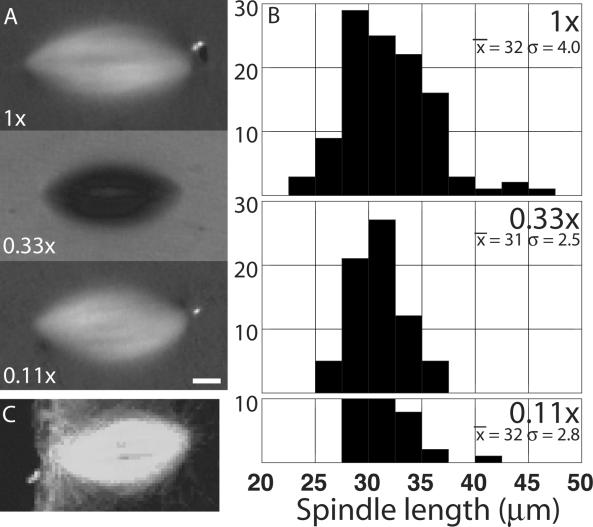
Metaphase spindles assemble to a steady state in length by mechanisms that involve microtubule dynamics and motor proteins, but they are incompletely understood. We found that Xenopus extract spindles recapitulate the length of egg meiosis II spindles, by using mechanisms intrinsic to the spindle. To probe these mechanisms, we perturbed microtubule polymerization dynamics and opposed motor proteins and measured effects on spindle morphology and dynamics. Microtubules were stabilized by hexylene glycol and inhibition of the catastrophe factor mitotic centromere-associated kinesin (MCAK) (a kinesin 13, previously called XKCM) and destabilized by depolymerizing drugs. The opposed motors Eg5 and dynein were inhibited separately and together. Our results are consistent with important roles for polymerization dynamics in regulating spindle length, and for opposed motors in regulating the relative stability of bipolar versus monopolar organization. The response to microtubule destabilization suggests that an unidentified tensile element acts in parallel with these conventional factors, generating spindle shortening force.
Figures








References
-
- Cassimeris, L., Rieder, C. L., Rupp, G., and Salmon, E. D. (1990). Stability of microtubule attachment to metaphase kinetochores in PtK1 cells. J. Cell Sci. 96, 9-15. - PubMed
-
- Cha, B. J., Error, B., and Gard, D. L. (1998). XMAP230 is required for the assembly and organization of acetylated microtubules and spindles in Xenopus oocytes and eggs. J. Cell Sci. 111, 2315-2327. - PubMed
-
- Chang, P., Jacobsen, M. K., and Mitchison, T. J. (2004). Poly(ADP-ribose) is required for spindle assembly and structure. Nature 432, 645-649. - PubMed
-
- Desai, A., Murray, A., Mitchison, T. J., and Walczak, C. E. (1999a). The use of Xenopus egg extracts to study mitotic spindle assembly and function in vitro. Methods Cell Biol. 61, 385-412. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
