

Chiral mutagenesis of insulin. Foldability and function are inversely regulated by a stereospecific switch in the B chain
- PMID: 15794637
- PMCID: PMC3845378
- DOI: 10.1021/bi048025o
Chiral mutagenesis of insulin. Foldability and function are inversely regulated by a stereospecific switch in the B chain
Abstract
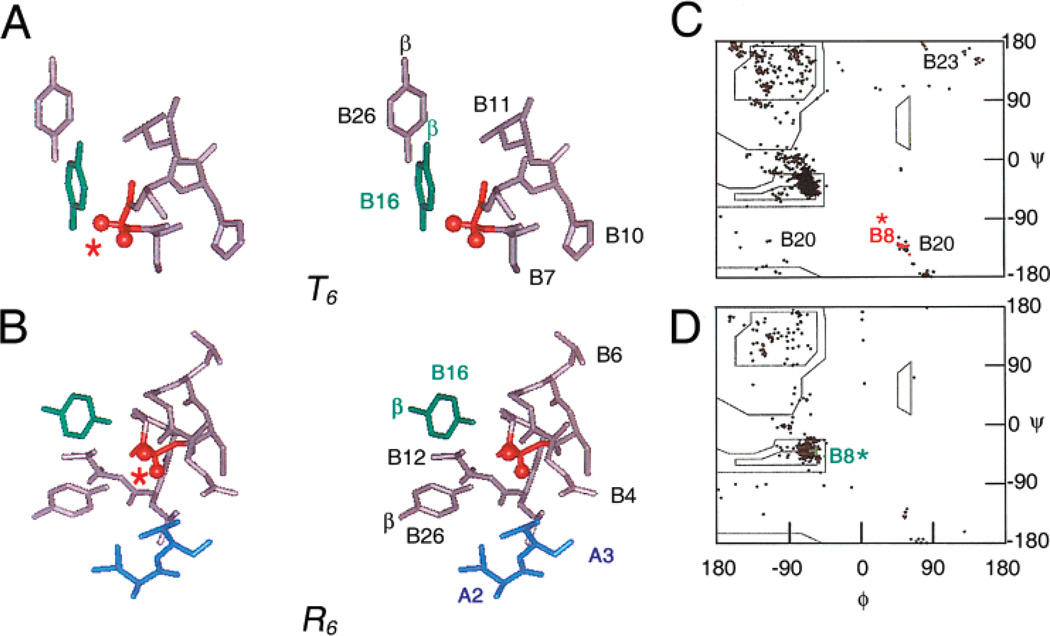
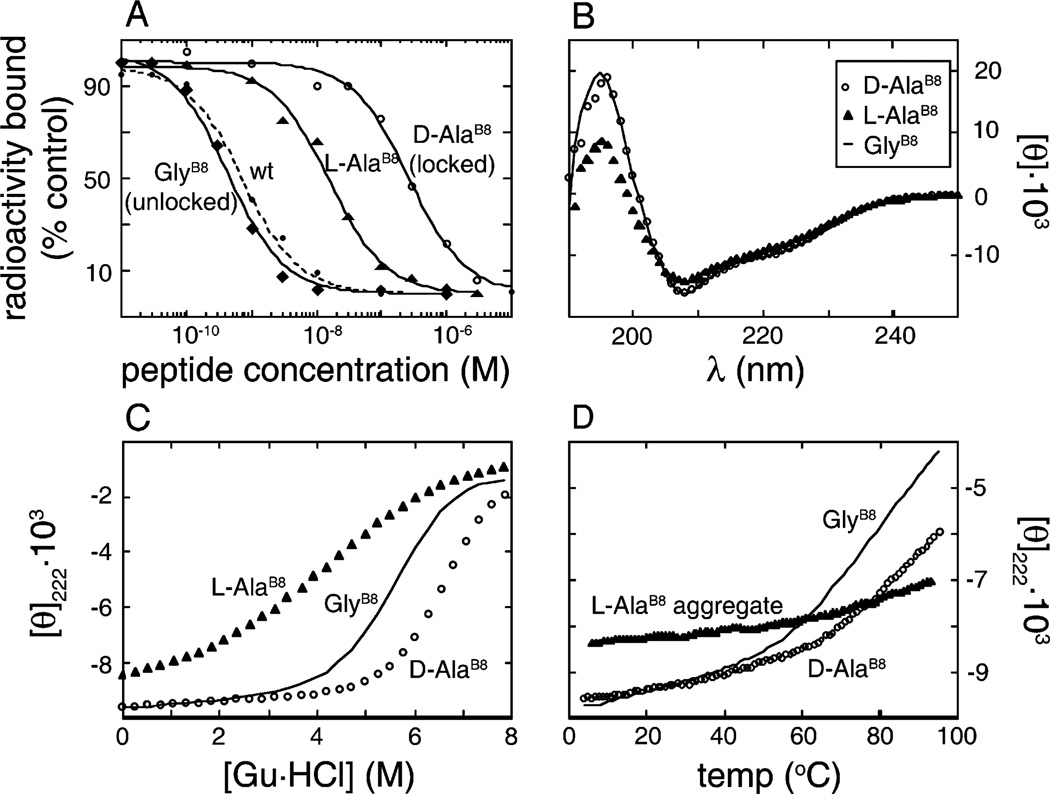
How insulin binds to its receptor is unknown despite decades of investigation. Here, we employ chiral mutagenesis-comparison of corresponding d and l amino acid substitutions in the hormone-to define a structural switch between folding-competent and active conformations. Our strategy is motivated by the T --> R transition, an allosteric feature of zinc-hexamer assembly in which an invariant glycine in the B chain changes conformations. In the classical T state, Gly(B8) lies within a beta-turn and exhibits a positive phi angle (like a d amino acid); in the alternative R state, Gly(B8) is part of an alpha-helix and exhibits a negative phi angle (like an l amino acid). Respective B chain libraries containing mixtures of d or l substitutions at B8 exhibit a stereospecific perturbation of insulin chain combination: l amino acids impede native disulfide pairing, whereas diverse d substitutions are well-tolerated. Strikingly, d substitutions at B8 enhance both synthetic yield and thermodynamic stability but markedly impair biological activity. The NMR structure of such an inactive analogue (as an engineered T-like monomer) is essentially identical to that of native insulin. By contrast, l analogues exhibit impaired folding and stability. Although synthetic yields are very low, such analogues can be highly active. Despite the profound differences between the foldabilities of d and l analogues, crystallization trials suggest that on protein assembly substitutions of either class can be accommodated within classical T or R states. Comparison between such diastereomeric analogues thus implies that the T state represents an inactive but folding-competent conformation. We propose that within folding intermediates the sign of the B8 phi angle exerts kinetic control in a rugged landscape to distinguish between trajectories associated with productive disulfide pairing (positive T-like values) or off-pathway events (negative R-like values). We further propose that the crystallographic T -->R transition in part recapitulates how the conformation of an insulin monomer changes on receptor binding. At the very least the ostensibly unrelated processes of disulfide pairing, allosteric assembly, and receptor binding appear to utilize the same residue as a structural switch; an "ambidextrous" glycine unhindered by the chiral restrictions of the Ramachandran plane. We speculate that this switch operates to protect insulin-and the beta-cell-from protein misfolding.
Figures







Similar articles
-
Toward the active conformation of insulin: stereospecific modulation of a structural switch in the B chain.J Biol Chem. 2006 Aug 25;281(34):24900-9. doi: 10.1074/jbc.M602691200. Epub 2006 Jun 8. J Biol Chem. 2006. PMID: 16762918
-
Chiral mutagenesis of insulin. Contribution of the B20-B23 beta-turn to activity and stability.J Biol Chem. 2006 Aug 4;281(31):22386-22396. doi: 10.1074/jbc.M603547200. Epub 2006 Jun 2. J Biol Chem. 2006. PMID: 16751187
-
Crystal structure of allo-Ile(A2)-insulin, an inactive chiral analogue: implications for the mechanism of receptor binding.Biochemistry. 2003 Nov 11;42(44):12770-83. doi: 10.1021/bi034430o. Biochemistry. 2003. PMID: 14596591
-
Zinc-ligand interactions modulate assembly and stability of the insulin hexamer -- a review.Biometals. 2005 Aug;18(4):295-303. doi: 10.1007/s10534-005-3685-y. Biometals. 2005. PMID: 16158220 Review.
-
Insulin: a small protein with a long journey.Protein Cell. 2010 Jun;1(6):537-51. doi: 10.1007/s13238-010-0069-z. Protein Cell. 2010. PMID: 21204007 Free PMC article. Review.
Cited by
-
Insulin analog with additional disulfide bond has increased stability and preserved activity.Protein Sci. 2013 Mar;22(3):296-305. doi: 10.1002/pro.2211. Epub 2013 Jan 17. Protein Sci. 2013. PMID: 23281053 Free PMC article.
-
Structural Ensemble of the Insulin Monomer.Biochemistry. 2021 Oct 26;60(42):3125-3136. doi: 10.1021/acs.biochem.1c00583. Epub 2021 Oct 12. Biochemistry. 2021. PMID: 34637307 Free PMC article.
-
Distinct states of proinsulin misfolding in MIDY.Cell Mol Life Sci. 2021 Aug;78(16):6017-6031. doi: 10.1007/s00018-021-03871-1. Epub 2021 Jul 10. Cell Mol Life Sci. 2021. PMID: 34245311 Free PMC article.
-
Insight into the structural and biological relevance of the T/R transition of the N-terminus of the B-chain in human insulin.Biochemistry. 2014 Jun 3;53(21):3392-402. doi: 10.1021/bi500073z. Epub 2014 May 22. Biochemistry. 2014. PMID: 24819248 Free PMC article.
-
Crystal structure of a "nonfoldable" insulin: impaired folding efficiency despite native activity.J Biol Chem. 2009 Dec 11;284(50):35259-72. doi: 10.1074/jbc.M109.046888. Epub 2009 Oct 22. J Biol Chem. 2009. PMID: 19850922 Free PMC article.
References
-
- Baker EN, Blundell TL, Cutfield JF, Cutfield SM, Dodson EJ, Dodson GG, Hodgkin DM, Hubbard RE, Isaacs NW, Reynolds CD. The structure of 2Zn pig insulin crystals at 1.5 Å resolution. Philos. Trans. Royal Soc. London Ser. 1988;319:369–456. - PubMed
-
- De Meyts P, Whittaker J. Structural biology of insulin and IGF–I receptors: implications for drug design. Nat. Rev. Drug Discovery. 2002;1:769–783. - PubMed
-
- Derewenda U, Derewenda Z, Dodson EJ, Dodson GG, Bing X, Markussen J. X-ray analysis of the single chain B29-A1 peptide-linked insulin molecule. A completely inactive analogue. J. Mol. Biol. 1991;220:425–433. - PubMed
-
- Hua QX, Shoelson SE, Kochoyan M, Weiss MA. Receptor binding redefined by a structural switch in a mutant human insulin. Nature. 1991;354:238–241. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
