

Review
doi: 10.1128/MCB.25.8.2873-2884.2005.
Class II histone deacetylases: from sequence to function, regulation, and clinical implication
Affiliations
- PMID: 15798178
- PMCID: PMC1069616
- DOI: 10.1128/MCB.25.8.2873-2884.2005
Item in Clipboard
Review
Class II histone deacetylases: from sequence to function, regulation, and clinical implication
Mol Cell Biol.
2005 Apr.
No abstract available
Figures
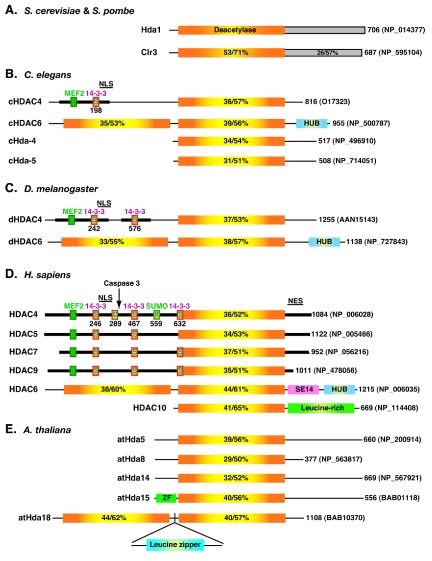
Domain organization of Hda1 and related proteins from fission yeast (A), worms (B), flies (C), humans (D), and plants (E). For each protein, only one isoform (usually the full-length isoform) is drawn, with its total number of residues and database accession number shown at the right. The deacetylase domain of Hda1 is depicted with a rectangle containing a yellow center flanked by orange areas. Similar rectangles are used to illustrate the deacetylase domains in Hda1-like proteins, with respective sequence identity/similarity to that of Hda1 listed. The C-terminal domains (shaded rectangles) of Hda1 and Clr3 are homologous. Dark bars, similar N-terminal domains and C-terminal tails of class IIa metazoan HDACs; small green boxes, MEF2-binding motifs; boxes labeled S (for serine), 14-3-3 binding motifs; boxes labeled D (for Asp289) and K (for Lys559), caspase cleavage and sumoylation sites on HDAC4, respectively. NES, nuclear export signal; SE14, SerGlu-containing tetradecapeptide repeats; ZF, zinc finger.
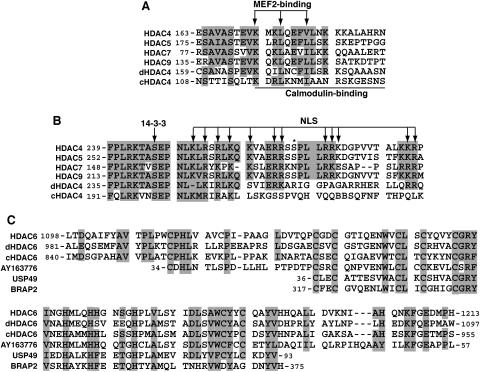
(A and B) Sequence alignment of the MEF2-binding motif (A) and the NLS (B) of class IIa HDACs. Residues invariant and highly conserved among at least four sequences are shaded. Arrows, residues known to be essential for the NLS function, 14-3-3 association, or MEF2 binding; solid line, region critical for Ca2+/calmodulin binding; asterisk, potential Dyrk1B phosphorylation site. (C) Sequence similarity between the HUB finger of HDAC6 and motifs in other proteins. Identical and highly conserved residues are shaded. Only the central region of the HUB finger is similar to USP49 and BRAP2. Residues invariant and highly conserved among at least 75% of the sequences are shaded.
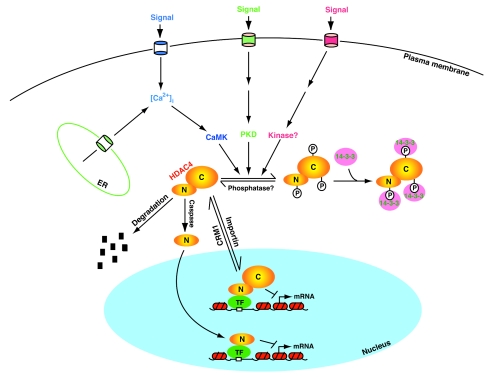
Cartoon depicting mechanisms involved in the regulation of HDAC4. It is actively imported and exported; the relative trafficking rate dictates its subcellular distribution. Cell signaling activates CaMK, PKD, and perhaps other kinases, thereby leading to site-specific phosphorylation of HDAC4 and association with 14-3-3 proteins. 14-3-3 binding then shifts the trafficking equilibrium of HDAC4 toward cytoplasmic accumulation. Unknown phosphatases dephosphorylate HDAC4 and dissociate it from 14-3-3 proteins for translocation to the nucleus, where it binds to sequence-specific transcription factors (TF) to repress transcription. The binding can be direct or mediated by a corepressor. Caspase 3 cleaves HDAC4 to generate the N-terminal (N) and C-terminal (C) fragments (Fig. 1D), with the former translocating to the nucleus as a transcriptional corepressor. HDAC4 may also be subject to proteosomal degradation. Dyrk1B and related kinases may phosphorylate HDAC4 and inhibit its nuclear import. Except for a few details, this model can be extended to other class IIa HDACs.

Sequence comparison of different α-tubulins. The peptides are flanked with the starting and ending positions, and residues invariant among at least five sequences are shaded. The sequence accession numbers are shown in parentheses at the right. Arrow, acetylation site. The acetylatable lysine residue is replaced by leucine in the only two α-tubulins found in budding yeast.
Similar articles
-
Deacetylase enzymes: biological functions and the use of small-molecule inhibitors.Chem Biol. 2002 Jan;9(1):3-16. doi: 10.1016/s1074-5521(02)00092-3. Chem Biol. 2002. PMID: 11841934 Review. No abstract available.
-
Identification of HDAC10, a novel class II human histone deacetylase containing a leucine-rich domain.Nucleic Acids Res. 2002 Mar 1;30(5):1114-23. doi: 10.1093/nar/30.5.1114. Nucleic Acids Res. 2002. PMID: 11861901 Free PMC article.
-
Histone deacetylase inhibitors and demethylating agents: clinical development of histone deacetylase inhibitors for cancer therapy.Cancer J. 2007 Jan-Feb;13(1):30-9. doi: 10.1097/PPO.0b013e31803c73cc. Cancer J. 2007. PMID: 17464244 Review.
-
Class IIa histone deacetylases: regulating the regulators.Oncogene. 2007 Aug 13;26(37):5450-67. doi: 10.1038/sj.onc.1210613. Oncogene. 2007. PMID: 17694086 Review.
-
HDAC8: a multifaceted target for therapeutic interventions.Trends Pharmacol Sci. 2015 Jul;36(7):481-92. doi: 10.1016/j.tips.2015.04.013. Epub 2015 May 23. Trends Pharmacol Sci. 2015. PMID: 26013035 Review.
Cited by
-
Transcriptional pathways and potential therapeutic targets in the regulation of Ncx1 expression in cardiac hypertrophy and failure.Adv Exp Med Biol. 2013;961:125-35. doi: 10.1007/978-1-4614-4756-6_11. Adv Exp Med Biol. 2013. PMID: 23224875 Free PMC article. Review.
-
Role of histone deacetylase inhibitors in non-neoplastic diseases.Heliyon. 2024 Jul 2;10(13):e33997. doi: 10.1016/j.heliyon.2024.e33997. eCollection 2024 Jul 15. Heliyon. 2024. PMID: 39071622 Free PMC article. Review.
-
Histone Deacetylases Exert Class-Specific Roles in Conditioning the Brain and Heart Against Acute Ischemic Injury.Front Neurol. 2015 Jun 30;6:145. doi: 10.3389/fneur.2015.00145. eCollection 2015. Front Neurol. 2015. PMID: 26175715 Free PMC article. Review.
-
Histone deacetylase inhibitors synergize p300 autoacetylation that regulates its transactivation activity and complex formation.Cancer Res. 2007 Mar 1;67(5):2256-2264. doi: 10.1158/0008-5472.CAN-06-3985. Cancer Res. 2007. PMID: 17332356 Free PMC article.
-
Effects of histone deacetylase inhibitors on HIF-1.Cell Cycle. 2006 Nov 1;5(21):2430-5. doi: 10.4161/cc.5.21.3409. Epub 2006 Sep 13. Cell Cycle. 2006. PMID: 17102633 Free PMC article. Review.
References
-
- Amann, J. M., J. Nip, D. K. Strom, B. Lutterbach, H. Harada, N. Lenny, J. R. Downing, S. Meyers, and S. W. Hiebert. 2001. ETO, a target of t(8;21) in acute leukemia, makes distinct contacts with multiple histone deacetylases and binds mSin3A through its oligomerization domain. Mol. Cell. Biol. 21:6470-6483. - PMC - PubMed
-
- Amerik, A. Y., S. J. Li, and M. Hochstrasser. 2000. Analysis of the deubiquitinating enzymes of the yeast Saccharomyces cerevisiae. Biol. Chem. 381:981-992. - PubMed
-
- Bakin, R. E., and M. O. Jung. 2004. Cytoplasmic sequestration of HDAC7 from mitochondrial and nuclear compartments upon initiation of apoptosis. J. Biol. Chem. 279:51218-51225. - PubMed
-
- Barlow, A. L., C. M. van Drunen, C. A. Johnson, S. Tweedie, A. Bird, and B. M. Turner. 2001. dSIR2 and dHDAC6: two novel, inhibitor-resistant deacetylases in Drosophila melanogaster. Exp. Cell Res. 265:90-103. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases