

Myocyte enhancer factor 2 activates promoter sequences of the human AbetaH-J-J locus, encoding aspartyl-beta-hydroxylase, junctin, and junctate
- PMID: 15798210
- PMCID: PMC1069596
- DOI: 10.1128/MCB.25.8.3261-3275.2005
Myocyte enhancer factor 2 activates promoter sequences of the human AbetaH-J-J locus, encoding aspartyl-beta-hydroxylase, junctin, and junctate
Abstract
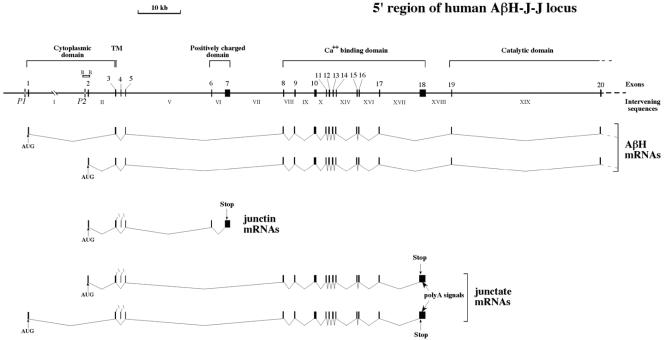
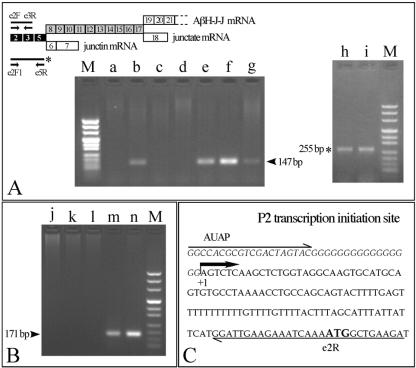
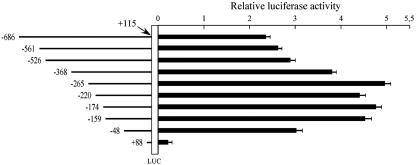
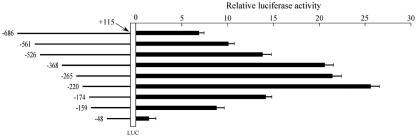
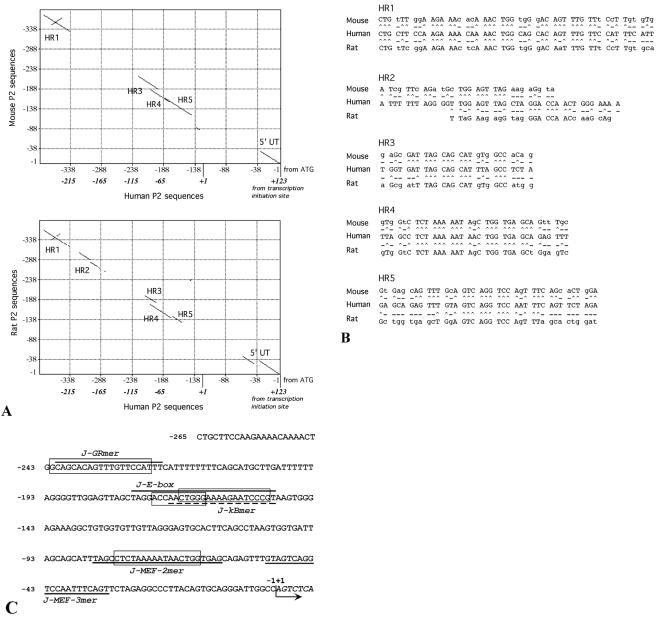
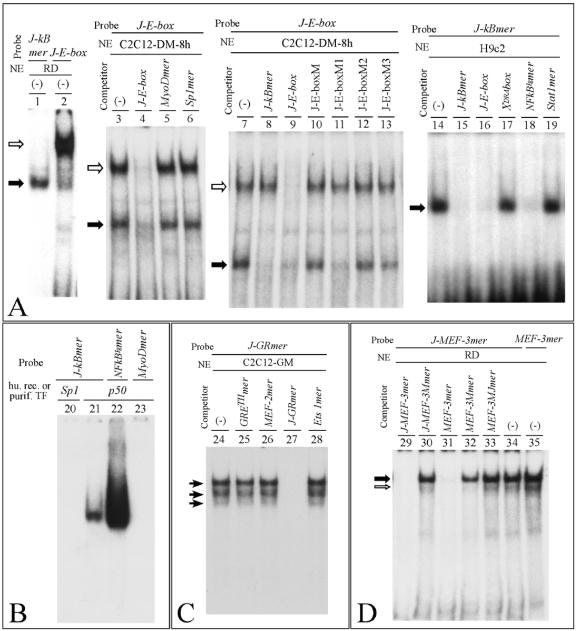
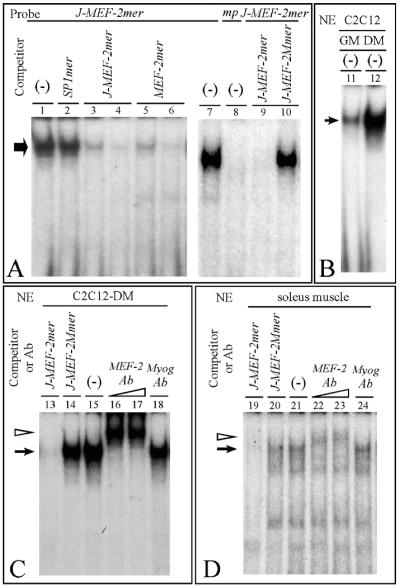
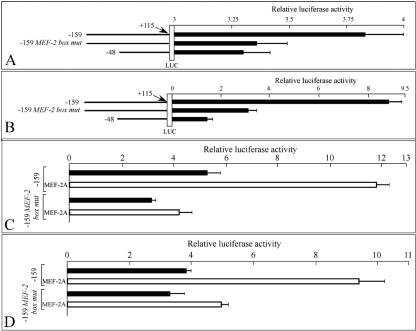
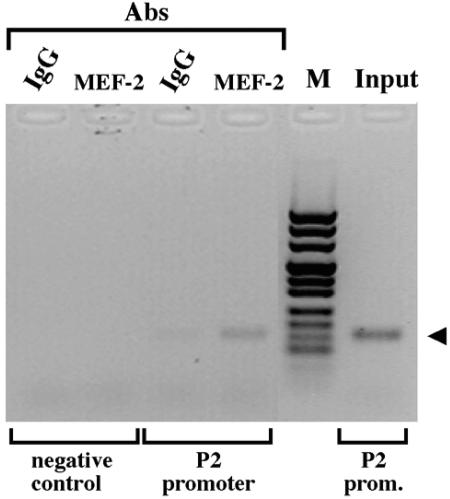
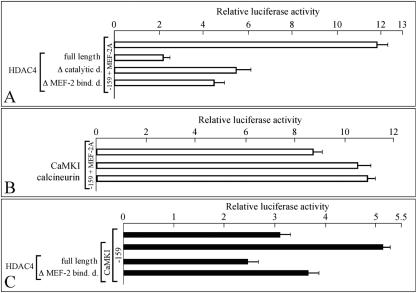
Alternative splicing of the locus AbetaH-J-J generates three functionally distinct proteins: an enzyme, AbetaH (aspartyl-beta-hydroxylase), a structural protein of the sarcoplasmic reticulum membrane (junctin), and an integral membrane calcium binding protein (junctate). Junctin and junctate are two important proteins involved in calcium regulation in eukaryotic cells. To understand the regulation of these two proteins, we identified and functionally characterized one of the two promoter sequences of the AbetaH-J-J locus. We demonstrate that the P2 promoter of the AbetaH-J-J locus contains (i) a minimal sequence localized within a region -159 bp from the transcription initiation site, which is sufficient to activate transcription of both mRNAs; (ii) sequences which bind known transcriptional factors such as those belonging to the myocyte enhancer factor 2 (MEF-2), MEF-3, and NF-kappaB protein families; and (iii) sequences bound by unknown proteins. The functional characterization of the minimal promoter in C2C12 cells and in the rat soleus muscle in vivo model indicates the existence of cis elements having positive and negative effects on transcription. In addition, our data demonstrate that in striated muscle cells the calcium-dependent transcription factor MEF-2 is crucial for the transcription activity directed by the P2 promoter. The transcription directed by the AbetaH-J-J P2 promoter is induced by high expression of MEF-2, further stimulated by calcineurin and Ca2+/calmodulin-dependent protein kinase I, and inhibited by histone deacetylase 4.
Figures

Similar articles
-
Multiple levels of control of the expression of the human A beta H-J-J locus encoding aspartyl-beta-hydroxylase, junctin, and junctate.Ann N Y Acad Sci. 2006 Dec;1091:184-90. doi: 10.1196/annals.1378.065. Ann N Y Acad Sci. 2006. PMID: 17341613 Review.
-
Transcriptional activity and Sp 1/3 transcription factor binding to the P1 promoter sequences of the human AbetaH-J-J locus.FEBS J. 2007 Sep;274(17):4476-90. doi: 10.1111/j.1742-4658.2007.05976.x. Epub 2007 Aug 6. FEBS J. 2007. PMID: 17681019
-
Molecular cloning, expression, functional characterization, chromosomal localization, and gene structure of junctate, a novel integral calcium binding protein of sarco(endo)plasmic reticulum membrane.J Biol Chem. 2000 Dec 15;275(50):39555-68. doi: 10.1074/jbc.M005473200. J Biol Chem. 2000. PMID: 11007777
-
Upstream stimulatory factors are involved in the P1 promoter directed transcription of the A beta H-J-J locus.BMC Mol Biol. 2008 Dec 16;9:110. doi: 10.1186/1471-2199-9-110. BMC Mol Biol. 2008. PMID: 19087304 Free PMC article.
-
Multiple functions of junctin and junctate, two distinct isoforms of aspartyl beta-hydroxylase.Biochem Biophys Res Commun. 2007 Oct 12;362(1):1-4. doi: 10.1016/j.bbrc.2007.07.166. Epub 2007 Aug 9. Biochem Biophys Res Commun. 2007. PMID: 17706594 Review.
Cited by
-
Aspartate/asparagine-β-hydroxylase: a high-throughput mass spectrometric assay for discovery of small molecule inhibitors.Sci Rep. 2020 May 26;10(1):8650. doi: 10.1038/s41598-020-65123-9. Sci Rep. 2020. PMID: 32457455 Free PMC article.
-
Regulatory roles of junctin in sarcoplasmic reticulum calcium cycling and myocardial function.Trends Cardiovasc Med. 2008 Jan;18(1):1-5. doi: 10.1016/j.tcm.2007.10.002. Trends Cardiovasc Med. 2008. PMID: 18206802 Free PMC article. Review.
-
Minor sarcoplasmic reticulum membrane components that modulate excitation-contraction coupling in striated muscles.J Physiol. 2009 Jul 1;587(Pt 13):3071-9. doi: 10.1113/jphysiol.2009.171876. Epub 2009 Apr 29. J Physiol. 2009. PMID: 19403606 Free PMC article. Review.
-
Vitamin K-dependent carboxylation regulates Ca2+ flux and adaptation to metabolic stress in β cells.Cell Rep. 2023 May 30;42(5):112500. doi: 10.1016/j.celrep.2023.112500. Epub 2023 May 11. Cell Rep. 2023. PMID: 37171959 Free PMC article.
-
Structure-function relationships and modifications of cardiac sarcoplasmic reticulum Ca2+-transport.Physiol Res. 2021 Dec 30;70(Suppl4):S443-S470. doi: 10.33549/physiolres.934805. Physiol Res. 2021. PMID: 35199536 Free PMC article. Review.
References
-
- Always, S. E., H. Degens, G. Krishnamurthy, and C. A. Smith. 2002. Potential role for Id myogenic repressors in apoptosis and attenuation of hypertrophy in muscles of aged rats. Am. J. Physiol. Cell Physiol. 283:C66-C76. - PubMed
-
- Ayoubi, T. A., and W. J. Van De Ven. 1996. Regulation of gene expression by alternative promoters. FASEB J. 10:453-460. - PubMed
-
- Black, B. L., J. F. Martin, and E. N. Olson. 1995. The mouse MRF4 promoter is trans-activated directly and indirectly by muscle-specific transcription factors. J. Biol. Chem. 270:2889-2892. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous