

Galectin-3 is a downstream regulator of matrix metalloproteinase-9 function during endochondral bone formation
- PMID: 15800063
- PMCID: PMC1142445
- DOI: 10.1091/mbc.e04-12-1119
Galectin-3 is a downstream regulator of matrix metalloproteinase-9 function during endochondral bone formation
Abstract
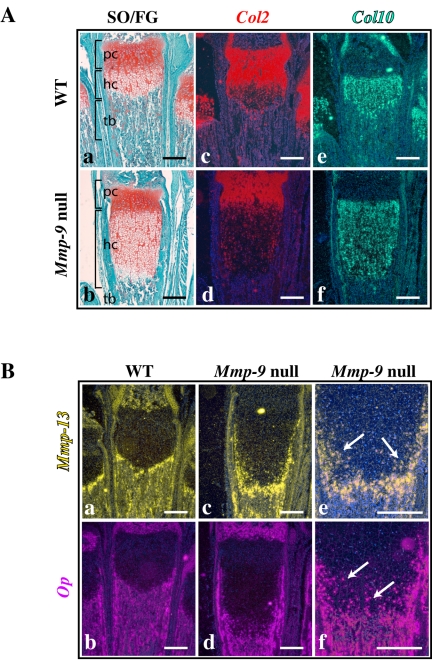
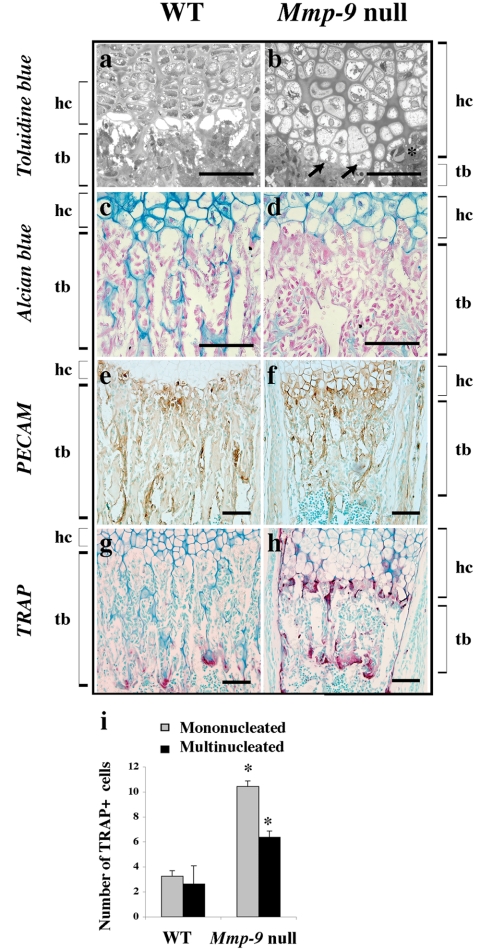
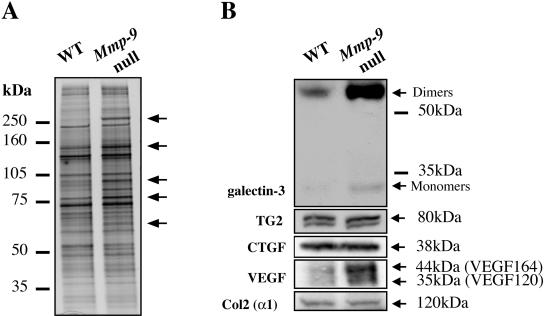
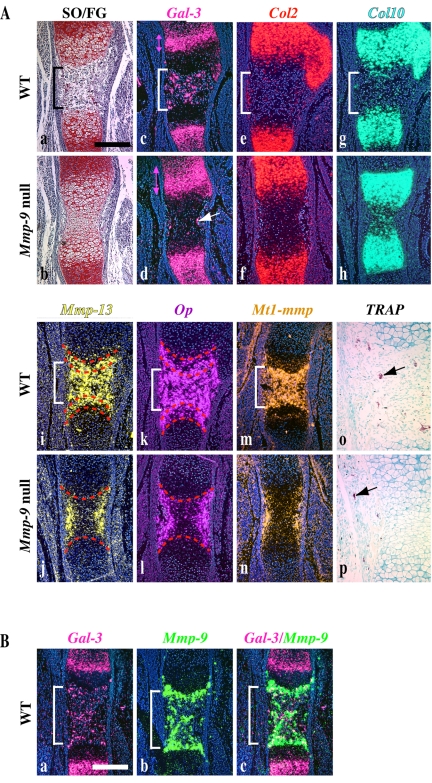
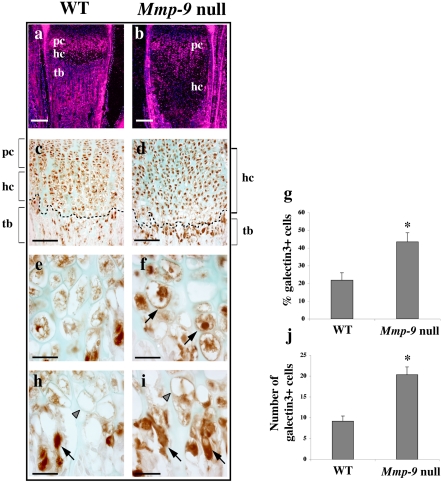
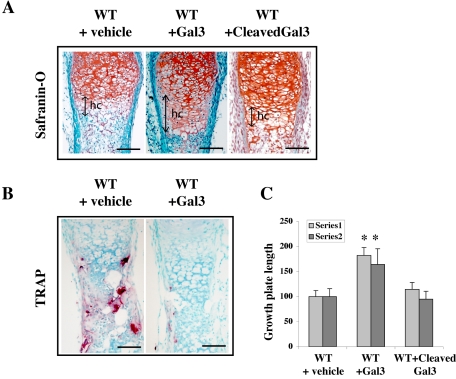
Endochondral bone formation is characterized by the progressive replacement of a cartilage anlagen by bone at the growth plate with a tight balance between the rates of chondrocyte proliferation, differentiation, and cell death. Deficiency of matrix metalloproteinase-9 (MMP-9) leads to an accumulation of late hypertrophic chondrocytes. We found that galectin-3, an in vitro substrate of MMP-9, accumulates in the late hypertrophic chondrocytes and their surrounding extracellular matrix in the expanded hypertrophic cartilage zone. Treatment of wild-type embryonic metatarsals in culture with full-length galectin-3, but not galectin-3 cleaved by MMP-9, mimicked the embryonic phenotype of Mmp-9 null mice, with an increased hypertrophic zone and decreased osteoclast recruitment. These results indicate that extracellular galectin-3 could be an endogenous substrate of MMP-9 that acts downstream to regulate hypertrophic chondrocyte death and osteoclast recruitment during endochondral bone formation. Thus, the disruption of growth plate homeostasis in Mmp-9 null mice links galectin-3 and MMP-9 in the regulation of the clearance of late chondrocytes through regulation of their terminal differentiation.
Figures
References
-
- Akahani, S., Nangia-Makker, P., Inohara, H., Kim, H. R., and Raz, A. (1997). Galectin-3, a novel antiapoptotic molecule with a functional BH1 (NWGR) domain of Bcl-2 family. Cancer Res. 57, 5272-5276. - PubMed
-
- Albrecht, U., Eichele, G., Helms, J. A., and Lu, H. (1997). Visualization of gene expression patterns by in situ hybridization. In: Molecular and Cellular Methods in Developmental Toxicology, ed. G. Daston, Boca Raton, FL: CRC Press, 23-48.
-
- Aubin, J. E., Gupta, A. K., Bhargava, U., and Turksen, K. (1996). Expression and regulation of galectin 3 in rat osteoblastic cells. J. Cell Physiol. 169, 468-480. - PubMed
-
- Barleon, B., Sozzani, S., Zhou, D., Weich, H. A., Mantovani, A., and Marme, D. (1996). Migration of human monocytes in response to vascular endothelial growth factor (VEGF) is mediated via the VEGF receptor flt-1. Blood 87, 3336-3343. - PubMed
-
- Barondes, S. H. et al. (1994). Galectins: a family of animal beta-galactoside-binding lectins. Cell 76, 597-598. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
