

Origins of host-specific populations of the blast pathogen Magnaporthe oryzae in crop domestication with subsequent expansion of pandemic clones on rice and weeds of rice
- PMID: 15802503
- PMCID: PMC1450392
- DOI: 10.1534/genetics.105.041780
Origins of host-specific populations of the blast pathogen Magnaporthe oryzae in crop domestication with subsequent expansion of pandemic clones on rice and weeds of rice
Abstract
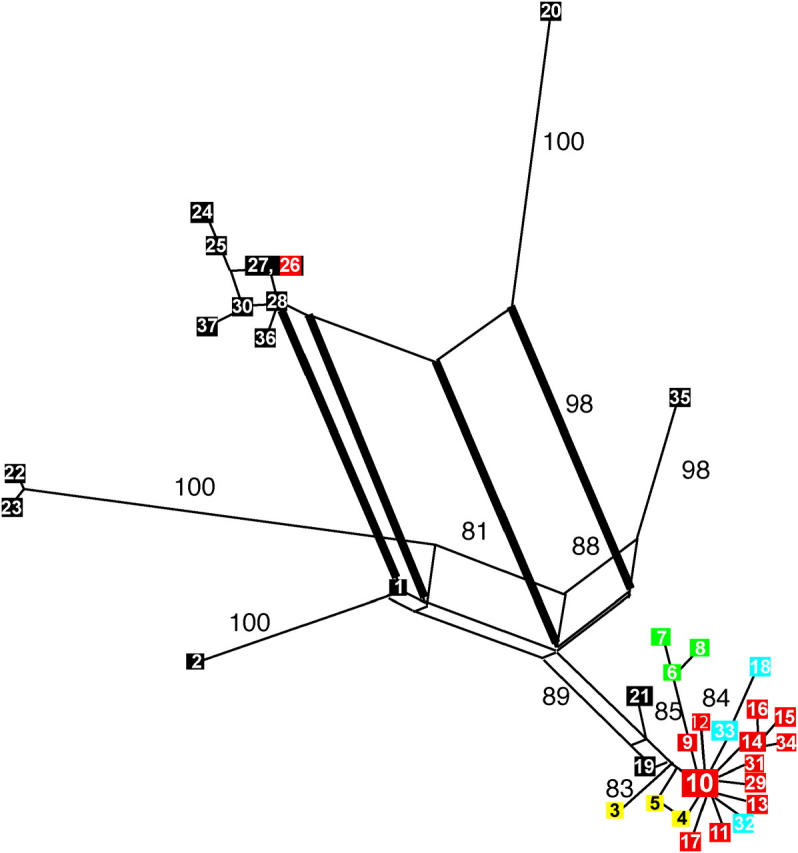
Rice, as a widely and intensively cultivated crop, should be a target for parasite host shifts and a source for shifts to co-occurring weeds. Magnaporthe oryzae, of the M. grisea species complex, is the most important fungal pathogen of rice, with a high degree of host specificity. On the basis of 10 loci from six of its seven linkage groups, 37 multilocus haplotypes among 497 isolates of M. oryzae from rice and other grasses were identified. Phylogenetic relationships among isolates from rice (Oryza sativa), millet (Setaria spp.), cutgrass (Leersia hexandra), and torpedo grass (Panicum repens) were predominantly tree like, consistent with a lack of recombination, but from other hosts were reticulate, consistent with recombination. The single origin of rice-infecting M. oryzae followed a host shift from a Setaria millet and was closely followed by additional shifts to weeds of rice, cutgrass, and torpedo grass. Two independent estimators of divergence time indicate that these host shifts predate the Green Revolution and could be associated with rice domestication. The rice-infecting lineage is characterized by high copy number of the transposable element MGR586 (Pot3) and, except in two haplotypes, by a loss of AVR-Co39. Both mating types have been retained in ancestral, well-distributed rice-infecting haplotypes 10 (mainly temperate) and 14 (mainly tropical), but only one mating type was recovered from several derived, geographically restricted haplotypes. There is evidence of a common origin of both ACE1 virulence genotypes in haplotype 14. Host-haplotype association is evidenced by low pathogenicity on hosts associated with other haplotypes.
Figures




References
-
- Antonovics, J., M. Hood and J. Partain, 2002. The ecology and genetics of a host-shift: Microbotryum as a model system. Am. Nat. 160: S40–S53. - PubMed
-
- Baker, R. H., and R. DeSalle, 1997. Multiple sources of character information and the phylogeny of Hawaiian drosophilids. Syst. Biol. 46: 654–673. - PubMed
-
- Baker, R. H., X. Yu and R. DeSalle, 1998. Assessing the relative contribution of molecular and morphological characters in simultaneous analysis trees. Mol. Phylogenet. Evol. 9: 427–436. - PubMed
-
- Bandelt, H. J., and A. W. Dress, 1992. Split decomposition: a new and useful approach to phylogenetic analysis of distance data. Mol. Phylogenet. Evol. 1: 242–252. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
