

Subdivision of large introns in Drosophila by recursive splicing at nonexonic elements
- PMID: 15802507
- PMCID: PMC1450422
- DOI: 10.1534/genetics.104.039701
Subdivision of large introns in Drosophila by recursive splicing at nonexonic elements
Abstract
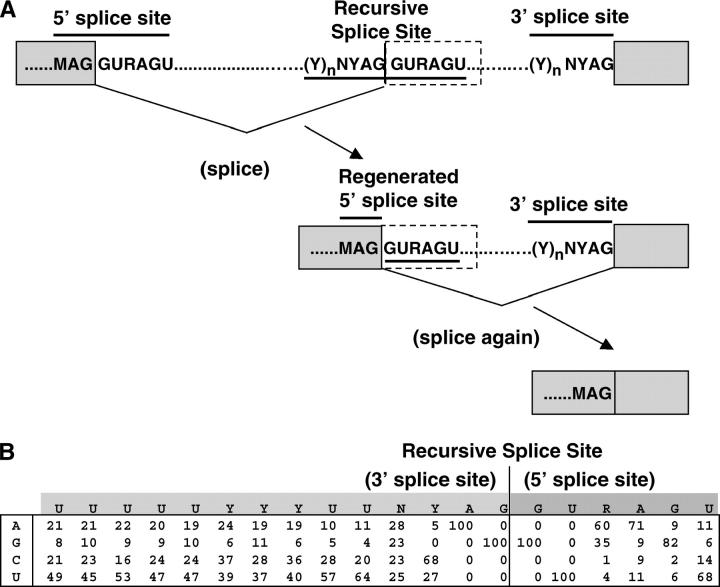
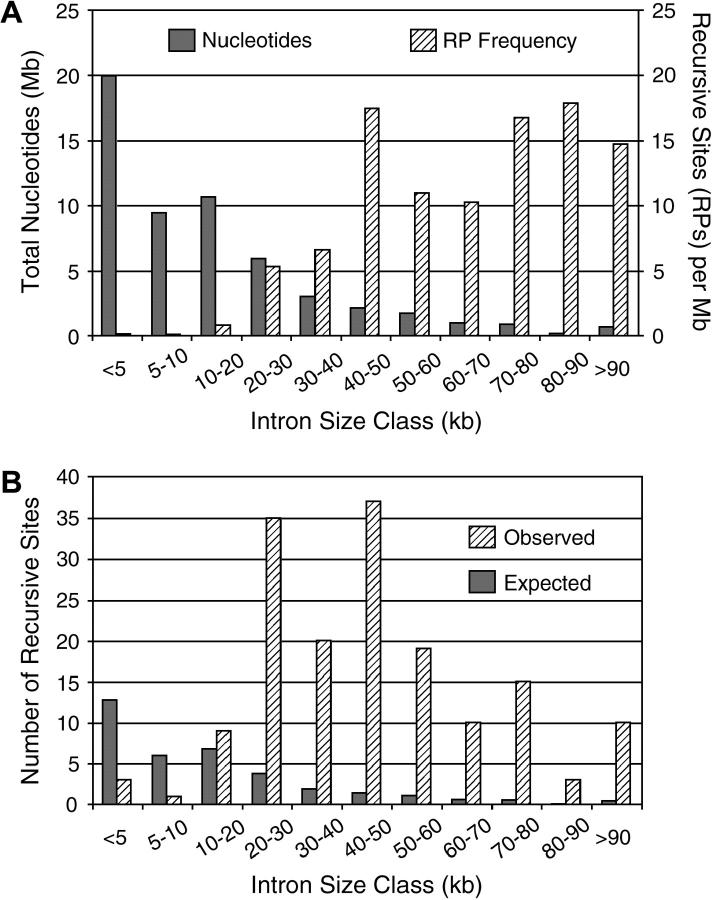
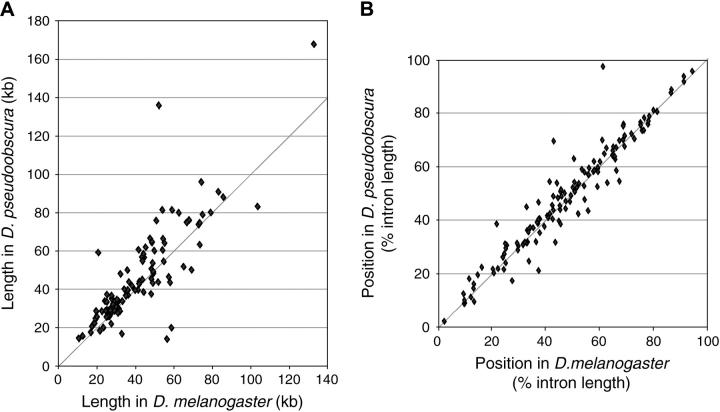
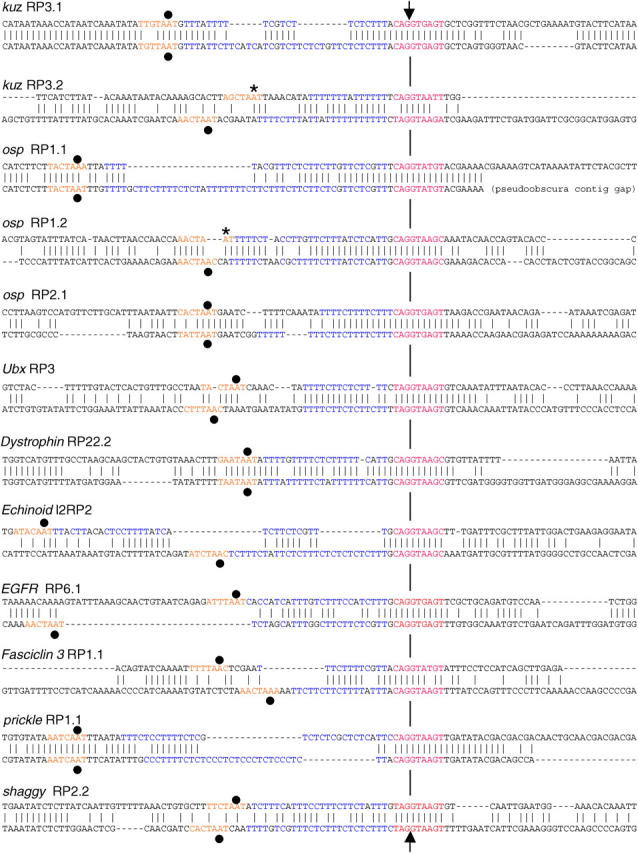
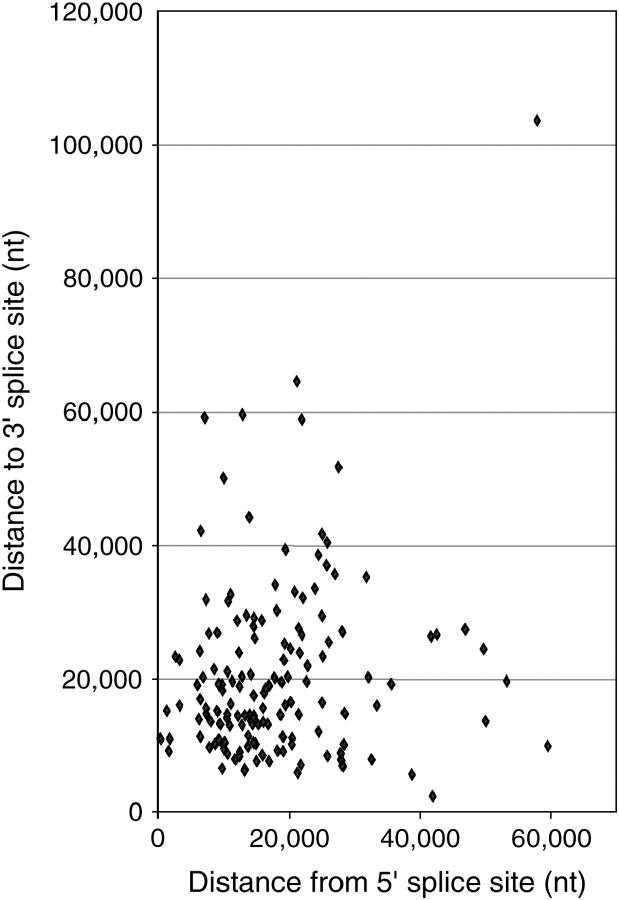
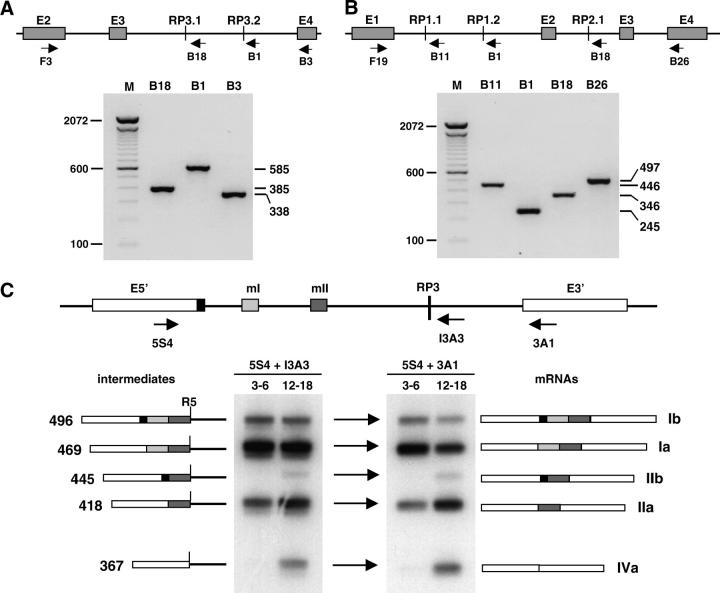
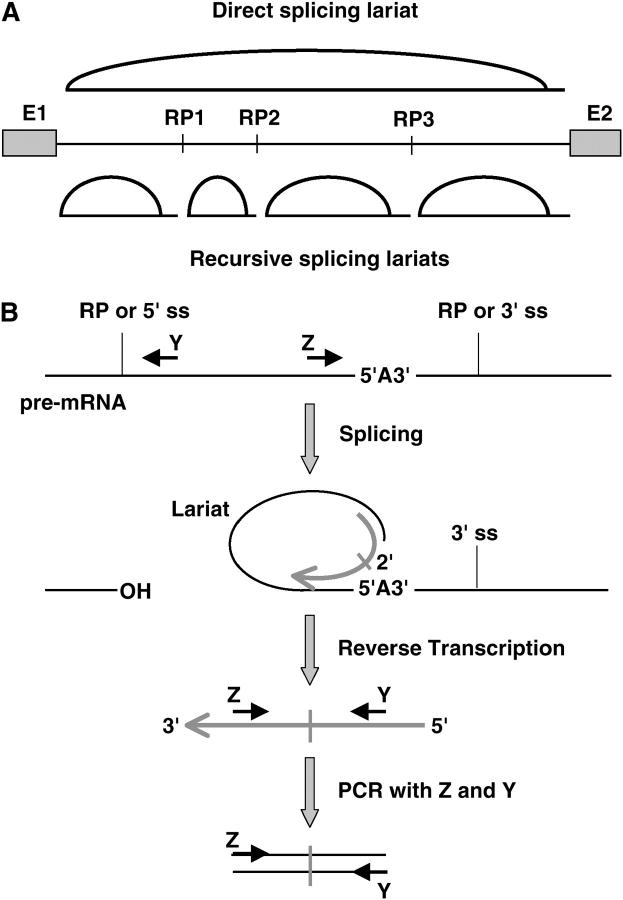
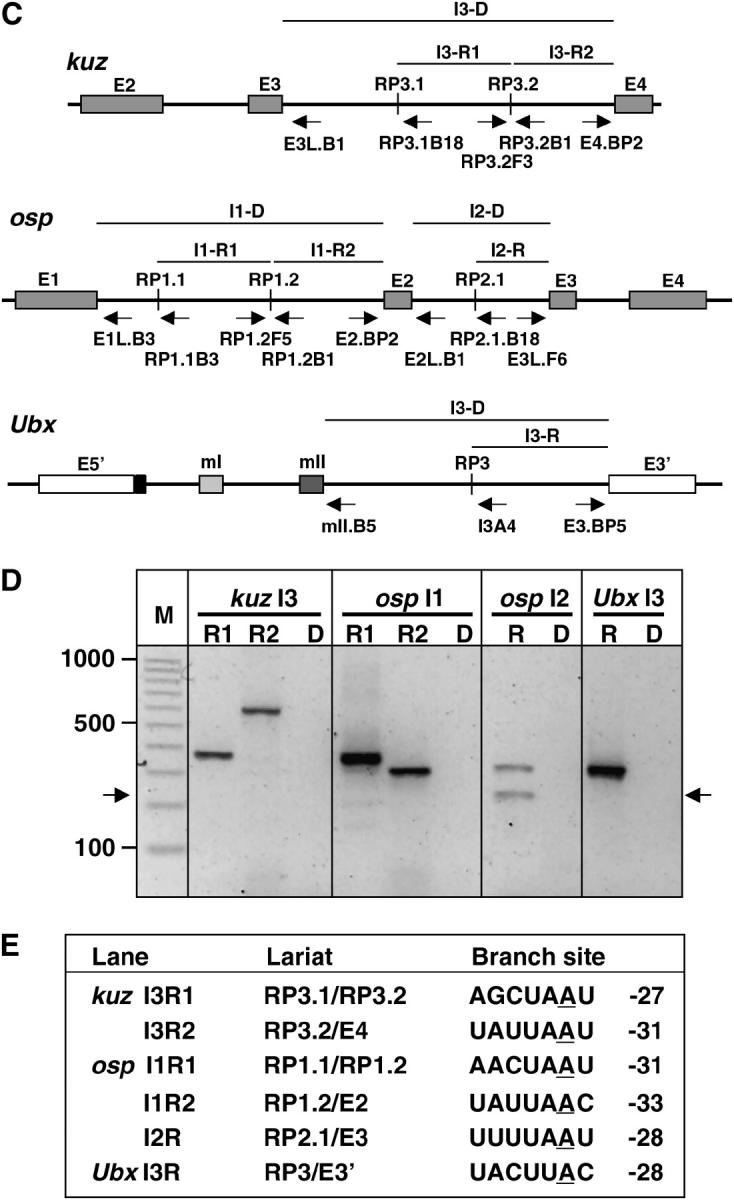
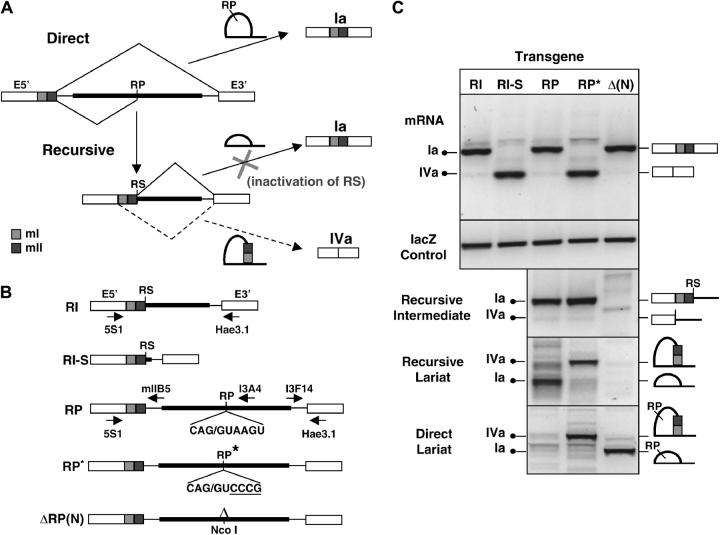
Many genes with important roles in development and disease contain exceptionally long introns, but special mechanisms for their expression have not been investigated. We present bioinformatic, phylogenetic, and experimental evidence in Drosophila for a mechanism that subdivides many large introns by recursive splicing at nonexonic elements and alternative exons. Recursive splice sites predicted with highly stringent criteria are found at much higher frequency than expected in the sense strands of introns >20 kb, but they are found only at the expected frequency on the antisense strands, and they are underrepresented within introns <10 kb. The predicted sites in long introns are highly conserved between Drosophila melanogaster and Drosophila pseudoobscura, despite extensive divergence of other sequences within the same introns. These patterns of enrichment and conservation indicate that recursive splice sites are advantageous in the context of long introns. Experimental analyses of in vivo processing intermediates and lariat products from four large introns in the unrelated genes kuzbanian, outspread, and Ultrabithorax confirmed that these introns are removed by a series of recursive splicing steps using the predicted nonexonic sites. Mutation of nonexonic site RP3 within Ultrabithorax also confirmed that recursive splicing is the predominant processing pathway even with a shortened version of the intron. We discuss currently known and potential roles for recursive splicing.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
