

A mitogen-activated protein kinase pathway essential for mating and contributing to vegetative growth in Neurospora crassa
- PMID: 15802524
- PMCID: PMC1451179
- DOI: 10.1534/genetics.104.036772
A mitogen-activated protein kinase pathway essential for mating and contributing to vegetative growth in Neurospora crassa
Abstract
MAP kinases homologous to Saccharomyces cerevisiae Fus3p/Kss1p have been identified in plant pathogenic fungi and are required for pathogenicity and sexual reproduction. To better understand the role of MAP kinase signaling in Neurospora crassa, and to identify downstream target genes of the pathway, we isolated, cloned, and disrupted the FUS3 homolog mak-2. Ste12p is a transcription factor target of Fus3p that activates genes of the mating pathway in yeast, and we also characterized the N. crassa STE12 homolog pp-1. The mak-2 and pp-1 mutants have reduced growth rate, produce short aerial hyphae, and fail to develop protoperithecia. In addition, ascospores carrying null mutations of either gene are inviable. Subtractive cloning was used to isolate genes having reduced expression in the mak-2 mutant. Expression of some of these genes is protoperithecia specific and three of them are part of a gene cluster potentially involved in the production of a polyketide secondary metabolite. Microarray analysis was used to extend the analysis of gene expression in mak-2 and pp-1 mutants. The role of the MAP kinase pathway in both sexual and asexual development as well as secondary metabolism is consistent with the dual regulation of the mating process and pathogencity observed in fungal pathogens.
Figures
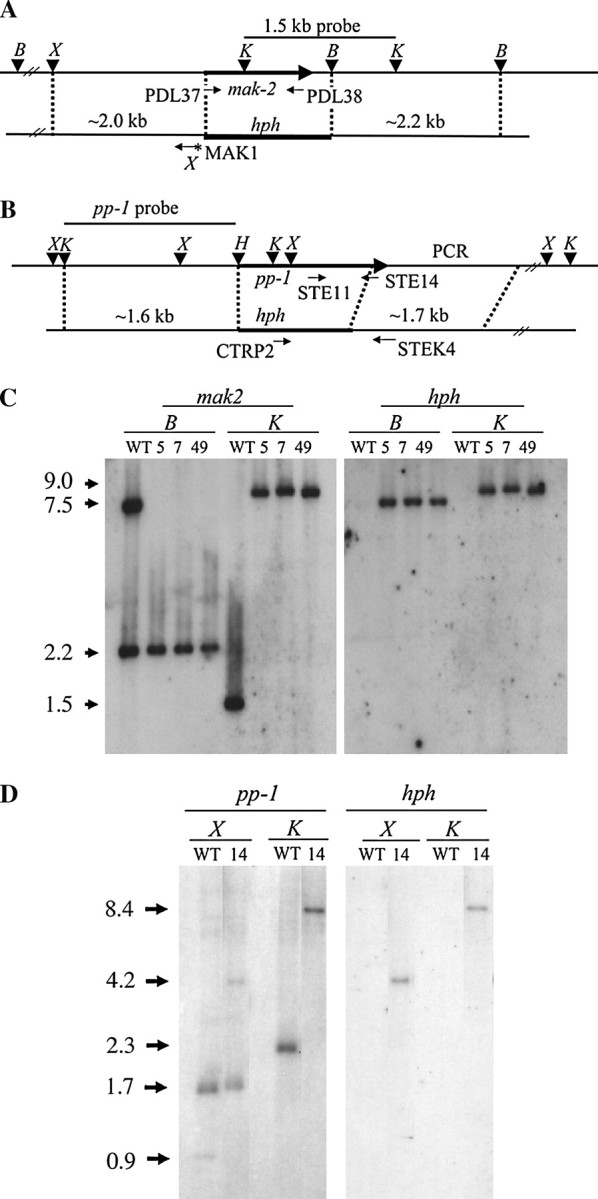
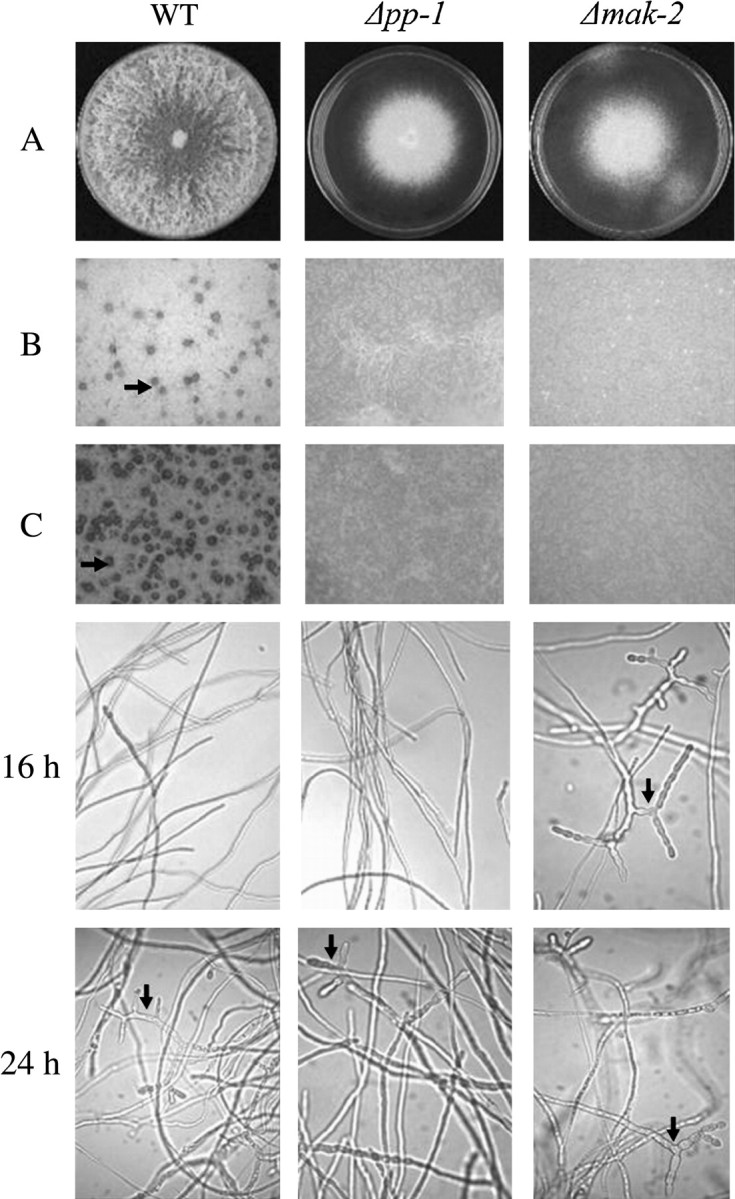

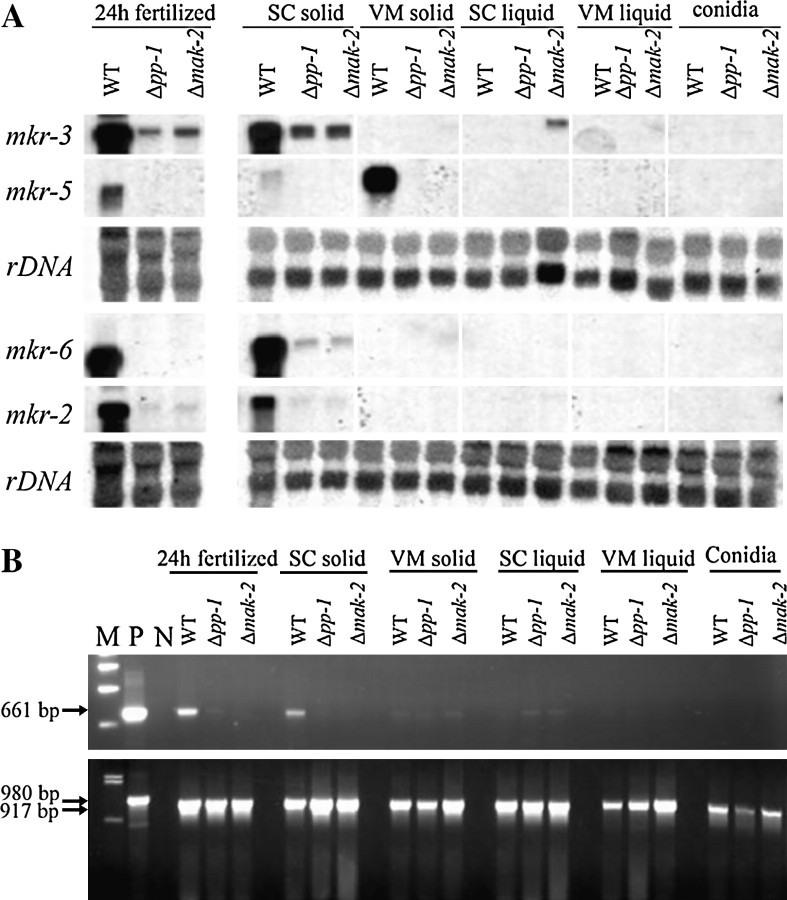

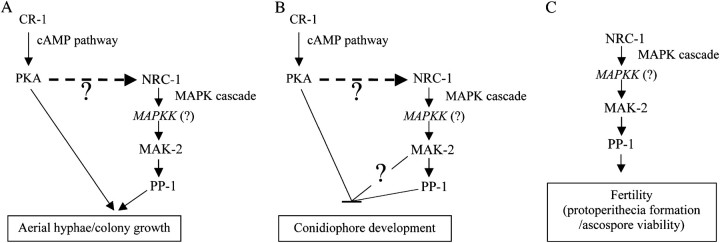
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
