

Common evolutionary origins of mechanosensitive ion channels in Archaea, Bacteria and cell-walled Eukarya
- PMID: 15803657
- PMCID: PMC2685541
- DOI: 10.1155/2002/419261
Common evolutionary origins of mechanosensitive ion channels in Archaea, Bacteria and cell-walled Eukarya
Abstract
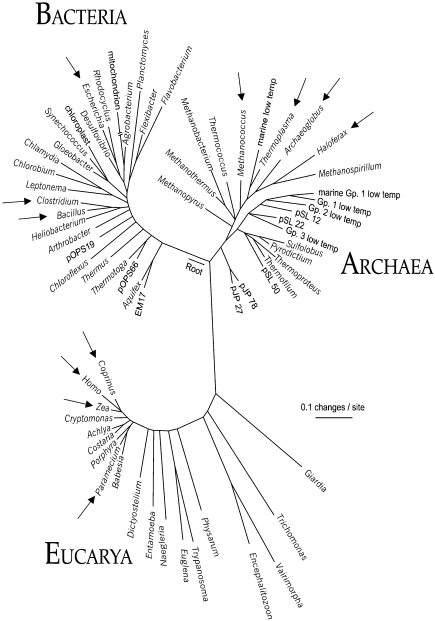
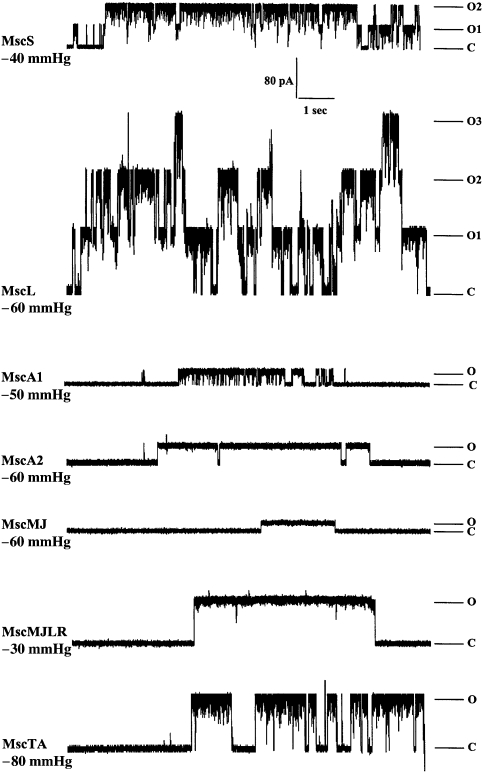
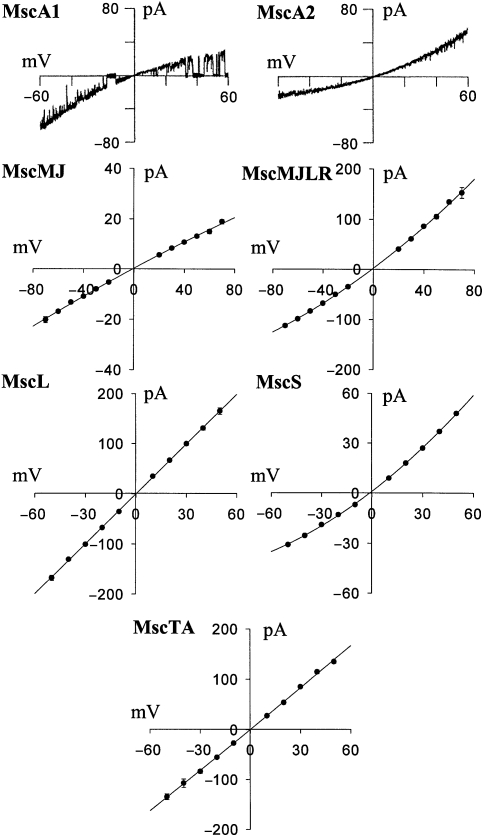
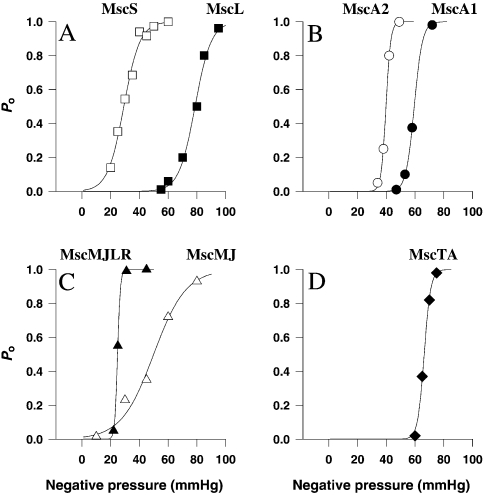
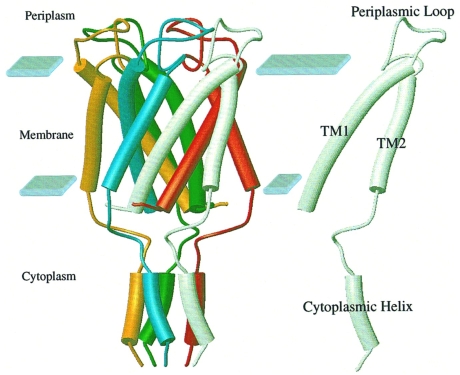
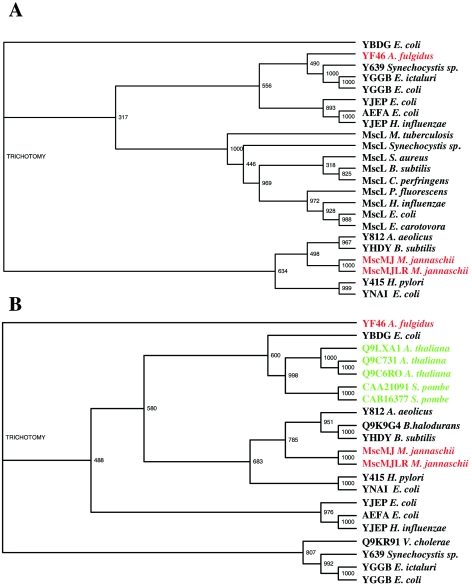
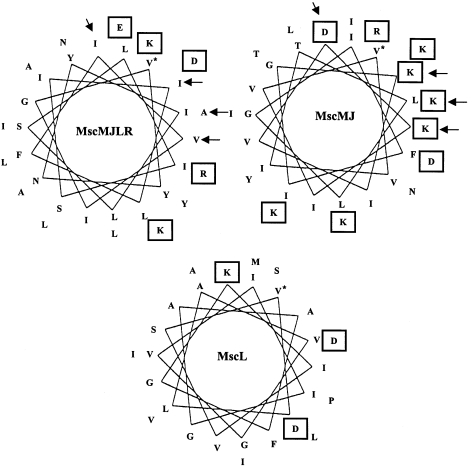
The ubiquity of mechanosensitive (MS) channels triggered a search for their functional homologs in Archaea. Archaeal MS channels were found to share a common ancestral origin with bacterial MS channels of large and small conductance, and sequence homology with several proteins that most likely function as MS ion channels in prokaryotic and eukaryotic cell-walled organisms. Although bacterial and archaeal MS channels differ in conductive and mechanosensitive properties, they share similar gating mechanisms triggered by mechanical force transmitted via the lipid bilayer. In this review, we suggest that MS channels of Archaea can bridge the evolutionary gap between bacterial and eukaryotic MS channels, and that MS channels of Bacteria, Archaea and cell-walled Eukarya may serve similar physiological functions and may have evolved to protect the fragile cellular membranes in these organisms from excessive dilation and rupture upon osmotic challenge.
Figures
References
-
- Ajouz B., Berrier C., Garrigues A., Besnard M., Ghazi A. Release of thioredoxin via mechanosensitive channel MscL during osmotic downshock of Escherichia coli cells. J. Biol. Chem. 1998;273:26670–26674. - PubMed
-
- Ajouz B., Berrier C., Besnard M., Martinac B., Ghazi A. Contributions of the different extramembraneous domains of the mechanosensitive ion channel MscL to its response to membrane tension. J. Biol. Chem. 2000;275:1015–1022. - PubMed
-
- Barinaga M. Molecular evolution. Archaea and eukaryotes grow closer. Science. 1994;264:1251. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
