

Regulation by external K+ in a maize inward shaker channel targets transport activity in the high concentration range
- PMID: 15805483
- PMCID: PMC1091772
- DOI: 10.1105/tpc.104.030551
Regulation by external K+ in a maize inward shaker channel targets transport activity in the high concentration range
Abstract
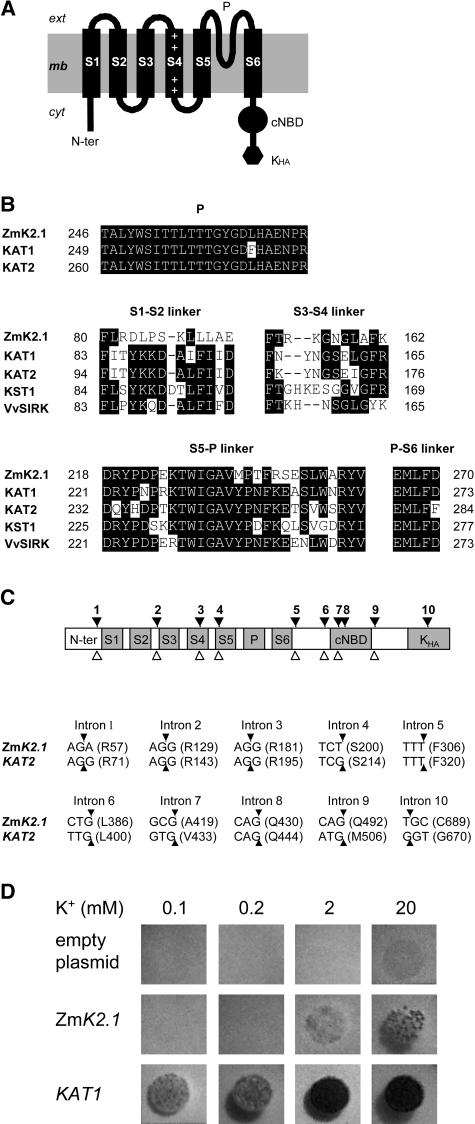
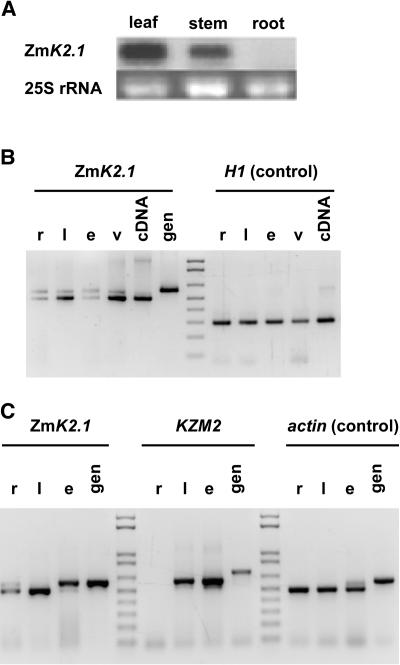
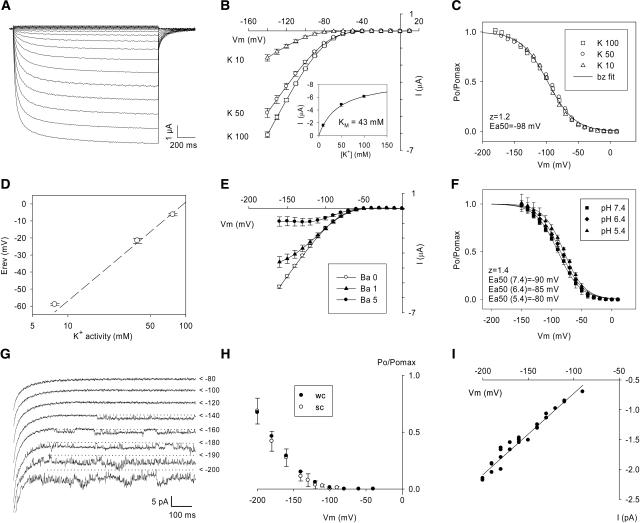
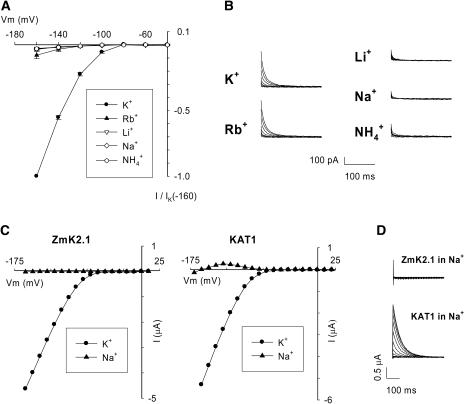
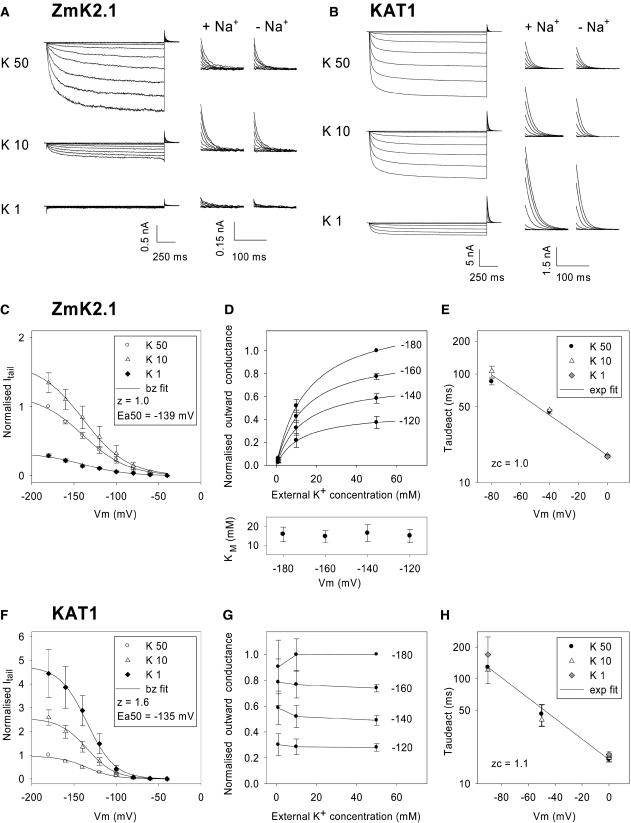
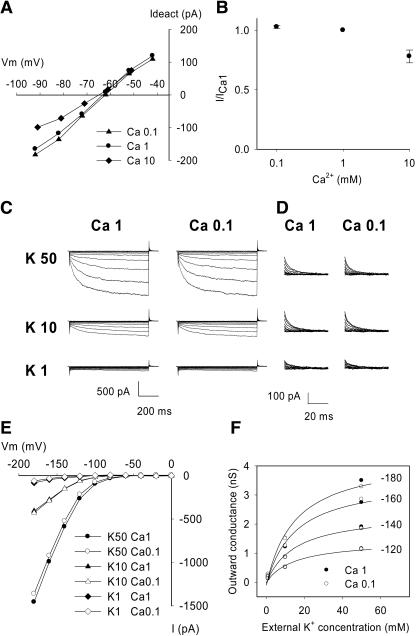
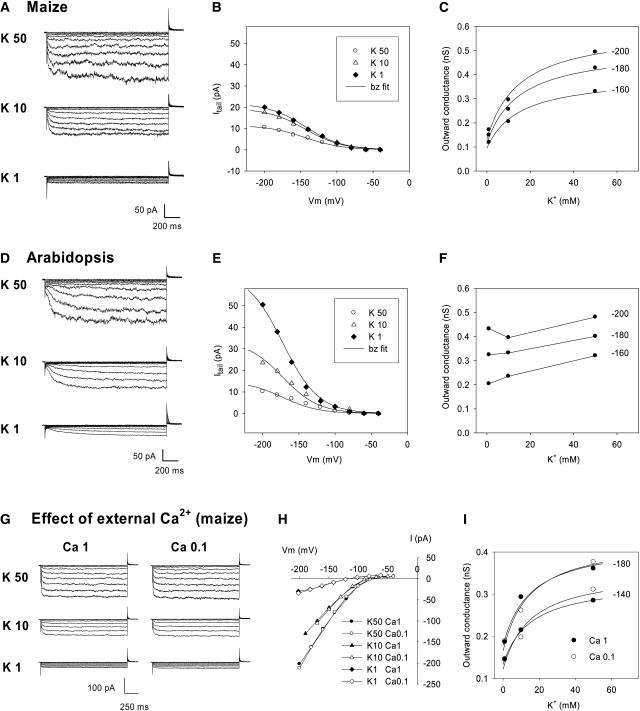
An inward Shaker K(+) channel identified in Zea mays (maize), ZmK2.1, displays strong regulation by external K(+) when expressed in Xenopus laevis (African clawed frog) oocytes or COS cells. ZmK2.1 is specifically activated by K(+) with an apparent K(m) close to 15 mM independent of the membrane hyperpolarization level. In the absence of K(+), ZmK2.1 appears to enter a nonconducting state. Thus, whatever the membrane potential, this maize channel cannot mediate K(+) influx in the submillimolar concentration range, unlike its relatives in Arabidopsis thaliana. Its expression is restricted to the shoots, the strongest signal (RT-PCR) being associated with vascular/bundle sheath strands. Based on sequence and gene structure, the closest relatives of ZmK2.1 in Arabidopsis are K(+) Arabidopsis Transporter 1 (KAT1) (expressed in guard cells) and KAT2 (expressed in guard cells and leaf phloem). Patch-clamp analyses of guard cell protoplasts reveal a higher functional diversity of K(+) channels in maize than in Arabidopsis. Channels endowed with regulation by external K(+) similar to that of ZmK2.1 (channel activity regulated by external K(+) with a K(m) close to 15 mM, regulation independent of external Ca(2+)) constitute a major component of the maize guard cell inward K(+) channel population. The presence of such channels in maize might reflect physiological traits of C4 and/or monocotyledonous plants.
Figures
Similar articles
-
The K+ channel KZM1 mediates potassium uptake into the phloem and guard cells of the C4 grass Zea mays.J Biol Chem. 2003 May 9;278(19):16973-81. doi: 10.1074/jbc.M212720200. Epub 2003 Feb 27. J Biol Chem. 2003. PMID: 12611901
-
AtKC1 is a general modulator of Arabidopsis inward Shaker channel activity.Plant J. 2011 Aug;67(4):570-82. doi: 10.1111/j.1365-313X.2011.04617.x. Epub 2011 Jul 11. Plant J. 2011. PMID: 21518051
-
The K+ channel KZM2 is involved in stomatal movement by modulating inward K+ currents in maize guard cells.Plant J. 2017 Nov;92(4):662-675. doi: 10.1111/tpj.13712. Epub 2017 Oct 20. Plant J. 2017. PMID: 28891257
-
Molecular insights into the structure and function of plant K(+) transport mechanisms.Biochim Biophys Acta. 2000 May 1;1465(1-2):127-39. doi: 10.1016/s0005-2736(00)00134-6. Biochim Biophys Acta. 2000. PMID: 10748250 Review.
-
Properties of shaker-type potassium channels in higher plants.J Membr Biol. 2006 Mar;210(1):1-19. doi: 10.1007/s00232-006-0856-x. Epub 2006 Jun 22. J Membr Biol. 2006. PMID: 16794778 Review.
Cited by
-
Speedy stomata of a C4 plant correlate with enhanced K+ channel gating.Plant Cell Environ. 2024 Mar;47(3):817-831. doi: 10.1111/pce.14775. Epub 2023 Nov 27. Plant Cell Environ. 2024. PMID: 38013592 Free PMC article.
-
Non-autonomous stomatal control by pavement cell turgor via the K+ channel subunit AtKC1.Plant Cell. 2022 Apr 26;34(5):2019-2037. doi: 10.1093/plcell/koac038. Plant Cell. 2022. PMID: 35157082 Free PMC article.
-
Saccharomyces cerevisiae as a Tool to Investigate Plant Potassium and Sodium Transporters.Int J Mol Sci. 2019 Apr 30;20(9):2133. doi: 10.3390/ijms20092133. Int J Mol Sci. 2019. PMID: 31052176 Free PMC article. Review.
-
Differential expression of K+ channels between guard cells and subsidiary cells within the maize stomatal complex.Planta. 2005 Dec;222(6):968-76. doi: 10.1007/s00425-005-0038-6. Epub 2005 Jul 15. Planta. 2005. PMID: 16021501
-
A procedure for localisation and electrophysiological characterisation of ion channels heterologously expressed in a plant context.Plant Methods. 2005 Dec 19;1:14. doi: 10.1186/1746-4811-1-14. Plant Methods. 2005. PMID: 16359560 Free PMC article.
References
-
- Assmann, S.M., and Wang, X.Q. (2001). From milliseconds to millions of years: Guard cells and environmental responses. Curr. Opin. Plant Biol. 4, 421–428. - PubMed
-
- Blatt, M.R. (1985). Extracellular potassium activity in attached leaves and its relation to stomatal function. J. Exp. Bot. 36, 240–251.
-
- Blatt, M.R. (1990). Potassium channel currents in intact stomatal guard cells: Rapid enhancement by abscisic acid. Planta 180, 445–455. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
