

Mutational comparison of the single-domained APOBEC3C and double-domained APOBEC3F/G anti-retroviral cytidine deaminases provides insight into their DNA target site specificities
- PMID: 15809227
- PMCID: PMC1074398
- DOI: 10.1093/nar/gki343
Mutational comparison of the single-domained APOBEC3C and double-domained APOBEC3F/G anti-retroviral cytidine deaminases provides insight into their DNA target site specificities
Abstract
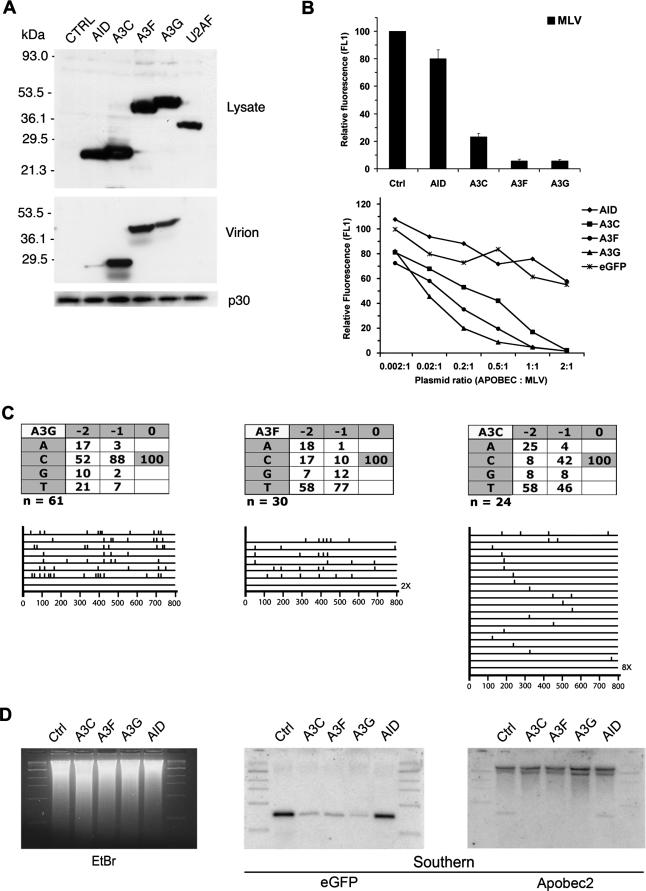
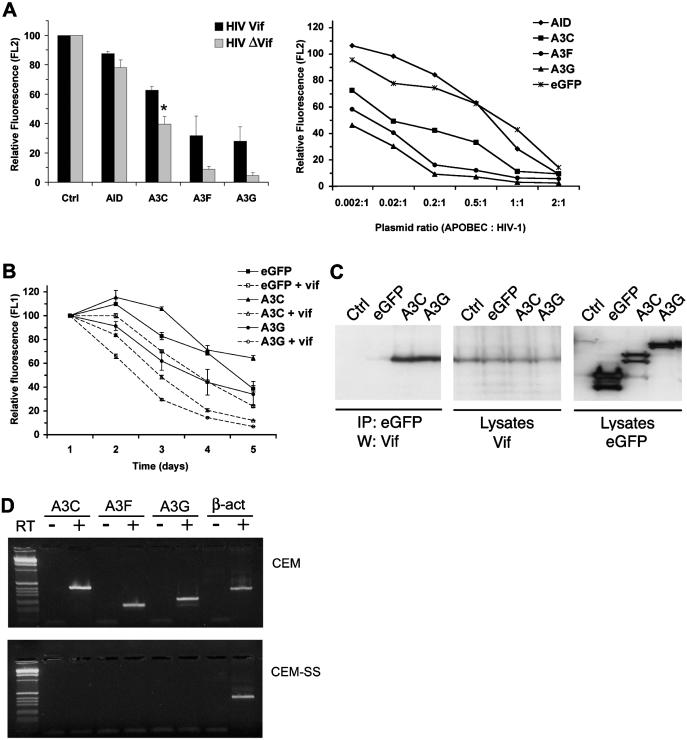
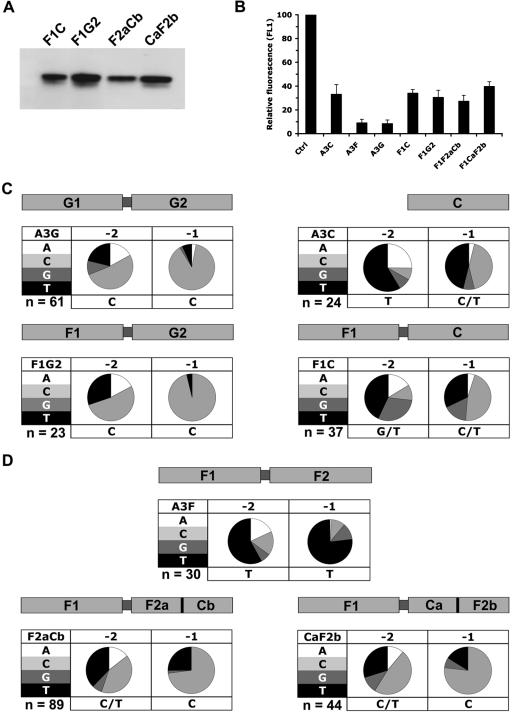
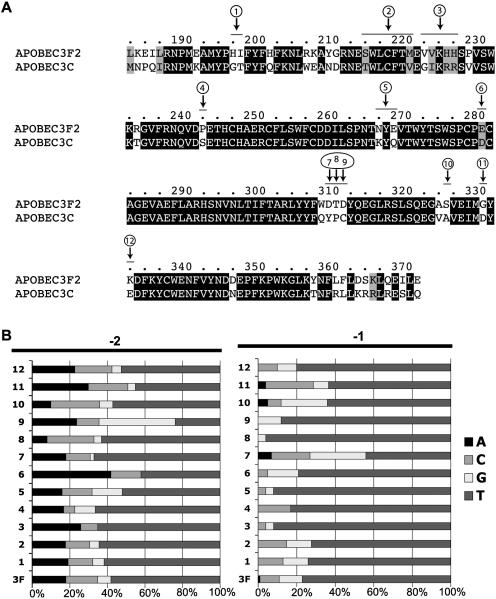
Human APOBEC3F and APOBEC3G are double-domained deaminases that can catalyze dC-->dU deamination in HIV-1 and MLV retroviral DNA replication intermediates, targeting T-C or C-C dinucleotides, respectively. HIV-1 antagonizes their action through its vif gene product, which has been shown (at least in the case of APOBEC3G) to interact with the N-terminal domain of the deaminase, triggering its degradation. Here, we compare APOBEC3F and APOBEC3G to APOBEC3C, a single-domained deaminase that can also act on both HIV-1 and MLV. We find that whereas APOBEC3C contains all the information necessary for both Vif-binding and cytidine deaminase activity in a single domain, it is the C-terminal domain of APOBEC3F and APOBEC3G that confer their target site specificity for cytidine deamination. We have exploited the fact that APOBEC3C, whilst highly homologous to the C-terminal domain of APOBEC3F, exhibits a distinct target site specificity (preferring Y-C dinucleotides) in order to identify residues in APOBEC3F that might affect its target site specificity. We find that this specificity can be altered by single amino acid substitutions at several distinct positions, suggesting that the strong dependence of APOBEC3-mediated deoxycytidine deamination on the 5'-flanking nucleotide is sensitive to relatively subtle changes in the APOBEC3 structure. The approach has allowed the isolation of APOBEC3 DNA mutators that exhibit novel target site preferences.
Figures
Similar articles
-
The retroviral hypermutation specificity of APOBEC3F and APOBEC3G is governed by the C-terminal DNA cytosine deaminase domain.J Biol Chem. 2005 Mar 25;280(12):10920-4. doi: 10.1074/jbc.M500382200. Epub 2005 Jan 12. J Biol Chem. 2005. PMID: 15647250
-
Biochemical differentiation of APOBEC3F and APOBEC3G proteins associated with HIV-1 life cycle.J Biol Chem. 2007 Jan 19;282(3):1585-94. doi: 10.1074/jbc.M610150200. Epub 2006 Dec 1. J Biol Chem. 2007. PMID: 17142455 Free PMC article.
-
APOBEC3F can inhibit the accumulation of HIV-1 reverse transcription products in the absence of hypermutation. Comparisons with APOBEC3G.J Biol Chem. 2007 Jan 26;282(4):2587-95. doi: 10.1074/jbc.M607298200. Epub 2006 Nov 22. J Biol Chem. 2007. PMID: 17121840
-
APOBEC deaminases as cellular antiviral factors: a novel natural host defense mechanism.Med Sci Monit. 2006 May;12(5):RA92-8. Med Sci Monit. 2006. PMID: 16641889 Review.
-
[Mechanisms for inhibition of retrovirus replication by APOBEC3 family].Uirusu. 2011 Jun;61(1):67-72. doi: 10.2222/jsv.61.67. Uirusu. 2011. PMID: 21972557 Review. Japanese.
Cited by
-
Correlation of APOBEC3 in tumor tissues with clinico-pathological features and survival from hepatocellular carcinoma after curative hepatectomy.Int J Clin Exp Med. 2015 May 15;8(5):7762-9. eCollection 2015. Int J Clin Exp Med. 2015. PMID: 26221327 Free PMC article.
-
Insights into the Structures and Multimeric Status of APOBEC Proteins Involved in Viral Restriction and Other Cellular Functions.Viruses. 2021 Mar 17;13(3):497. doi: 10.3390/v13030497. Viruses. 2021. PMID: 33802945 Free PMC article. Review.
-
Binding of RNA by APOBEC3G controls deamination-independent restriction of retroviruses.Nucleic Acids Res. 2013 Aug;41(15):7438-52. doi: 10.1093/nar/gkt527. Epub 2013 Jun 12. Nucleic Acids Res. 2013. PMID: 23761443 Free PMC article.
-
An ancient history of gene duplications, fusions and losses in the evolution of APOBEC3 mutators in mammals.BMC Evol Biol. 2012 May 28;12:71. doi: 10.1186/1471-2148-12-71. BMC Evol Biol. 2012. PMID: 22640020 Free PMC article.
-
All APOBEC3 family proteins differentially inhibit LINE-1 retrotransposition.Nucleic Acids Res. 2007;35(9):2955-64. doi: 10.1093/nar/gkm181. Epub 2007 Apr 16. Nucleic Acids Res. 2007. PMID: 17439959 Free PMC article.
References
-
- Sheehy A.M., Gaddis N.C., Choi J.D., Malim M.H. Isolation of a human gene that inhibits HIV-1 infection and is suppressed by the viral Vif protein. Nature. 2002;418:646–650. - PubMed
-
- Marin M., Rose K.M., Kozak S.L., Kabat D. HIV-1 Vif protein binds the editing enzyme APOBEC3G and induces its degradation. Nature Med. 2003;9:1398–1403. - PubMed
-
- Stopak K., de Noronha C., Yonemoto W., Greene W.C. HIV-1 Vif blocks the antiviral activity of APOBEC3G by impairing both its translation and intracellular stability. Mol. Cell. 2003;12:591–601. - PubMed
-
- Sheehy A.M., Gaddis N.C., Malim M.H. The antiretroviral enzyme APOBEC3G is degraded by the proteasome in response to HIV-1 Vif. Nature Med. 2003;9:1404–1407. - PubMed
-
- Yu X., Yu Y., Liu B., Luo K., Kong W., Mao P., Yu X.F. Induction of APOBEC3G ubiquitination and degradation by an HIV-1 Vif-Cul5-SCF complex. Science. 2003;302:1056–1060. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
