

NMDA receptors mediate olfactory learning and memory in Drosophila
- PMID: 15823532
- PMCID: PMC3045563
- DOI: 10.1016/j.cub.2005.02.059
NMDA receptors mediate olfactory learning and memory in Drosophila
Abstract
Background: Molecular and electrophysiological properties of NMDARs suggest that they may be the Hebbian "coincidence detectors" hypothesized to underlie associative learning. Because of the nonspecificity of drugs that modulate NMDAR function or the relatively chronic genetic manipulations of various NMDAR subunits from mammalian studies, conclusive evidence for such an acute role for NMDARs in adult behavioral plasticity, however, is lacking. Moreover, a role for NMDARs in memory consolidation remains controversial.
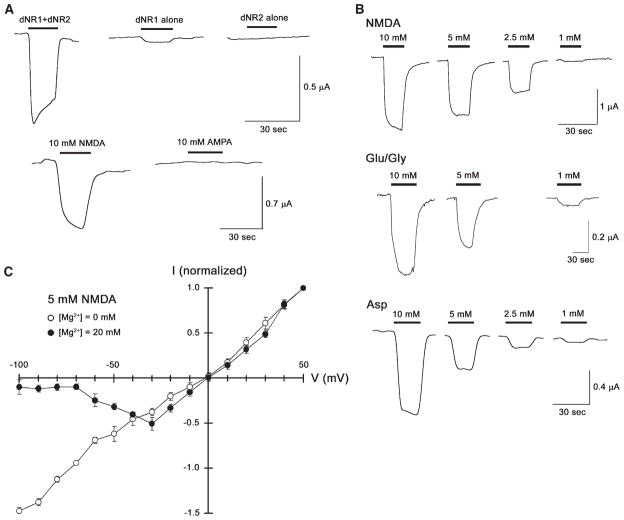
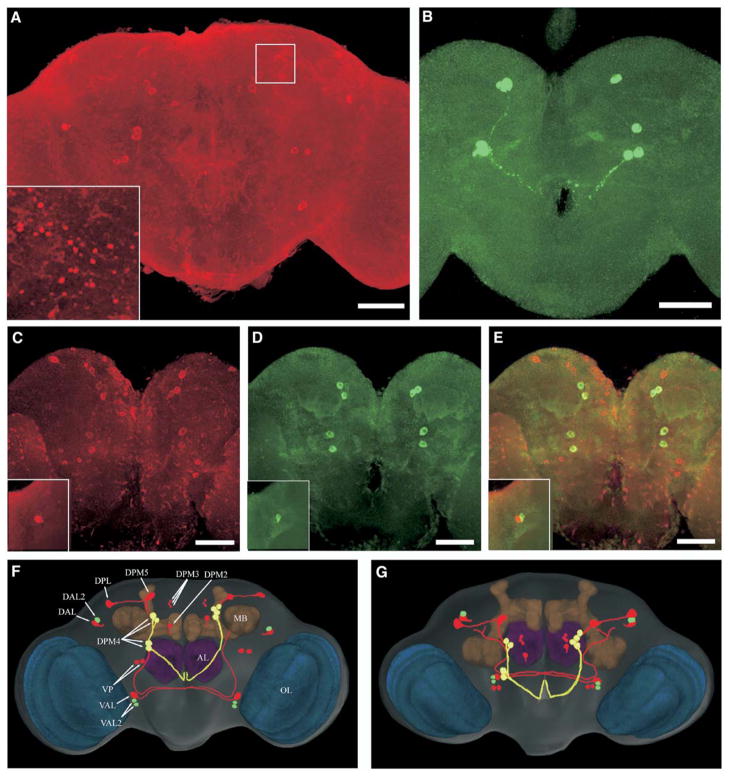
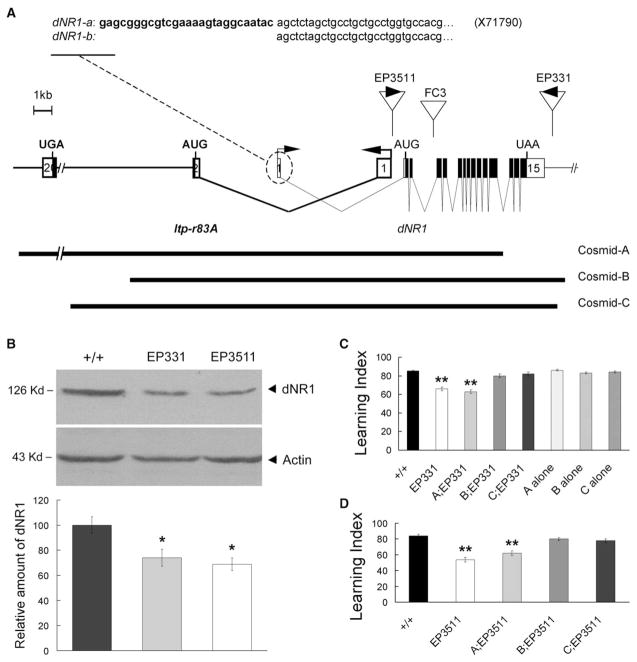
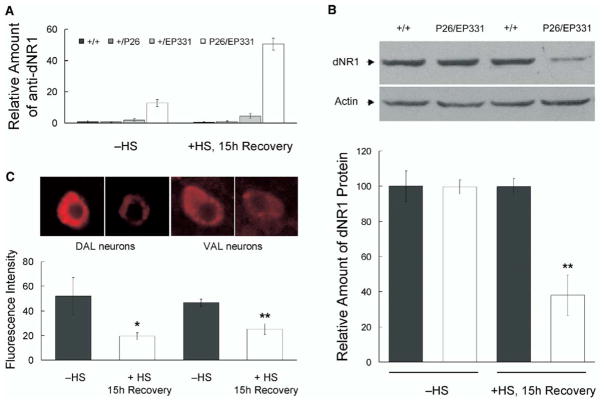
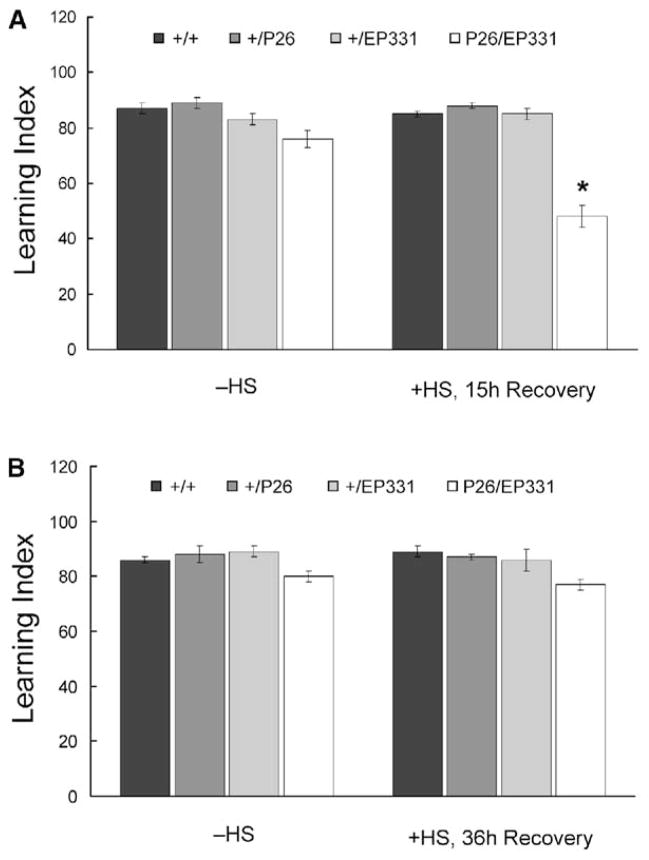
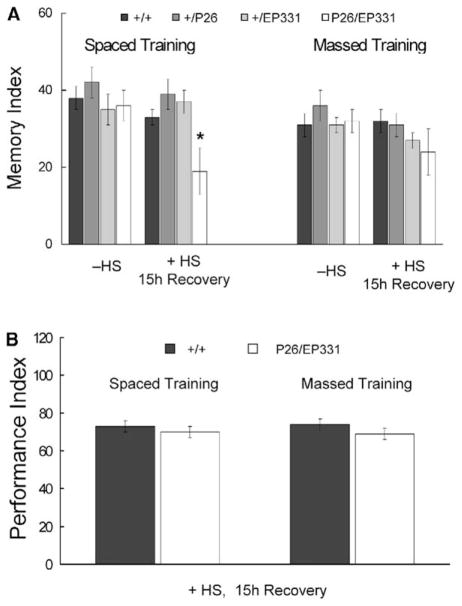
Results: The Drosophila genome encodes two NMDAR homologs, dNR1 and dNR2. When coexpressed in Xenopus oocytes or Drosophila S2 cells, dNR1 and dNR2 form functional NMDARs with several of the distinguishing molecular properties observed for vertebrate NMDARs, including voltage/Mg(2+)-dependent activation by glutamate. Both proteins are weakly expressed throughout the entire brain but show preferential expression in several neurons surrounding the dendritic region of the mushroom bodies. Hypomorphic mutations of the essential dNR1 gene disrupt olfactory learning, and this learning defect is rescued with wild-type transgenes. Importantly, we show that Pavlovian learning is disrupted in adults within 15 hr after transient induction of a dNR1 antisense RNA transgene. Extended training is sufficient to overcome this initial learning defect, but long-term memory (LTM) specifically is abolished under these training conditions.
Conclusions: Our study uses a combination of molecular-genetic tools to (1) generate genomic mutations of the dNR1 gene, (2) rescue the accompanying learning deficit with a dNR1+ transgene, and (3) rapidly and transiently knockdown dNR1+ expression in adults, thereby demonstrating an evolutionarily conserved role for the acute involvement of NMDARs in associative learning and memory.
Figures

References
-
- McBain CJ, Mayer ML. N-methyl-D-aspartic acid receptor structure and function. Physiol Rev. 1994;74:723–760. - PubMed
-
- Monaghan DT, Bridges RJ, Cotman CW. The excitatory amino acid receptors: their classes, pharmacology, and distinct properties in the function of the central nervous system. Annu Rev Pharmacol Toxicol. 1989;29:365–402. - PubMed
-
- Nowak L, Bregestovski P, Ascher P, Herbet A, Prochiantz A. Magnesium gates glutamate-activated channels in mouse central neurones. Nature. 1984;307:462–465. - PubMed
-
- Abbott LF, Nelson SB. Synaptic plasticity: taming the beast. Nat Neurosci. 2000;307(Suppl):1178–1183. - PubMed
-
- Debanne D, Daoudal G, Sourdet V, Russier M. Brain plasticity and ion channels. J Physiol (Paris) 2003;97:403–414. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
