

Late-onset motoneuron disease caused by a functionally modified AMPA receptor subunit
- PMID: 15827116
- PMCID: PMC556301
- DOI: 10.1073/pnas.0501316102
Late-onset motoneuron disease caused by a functionally modified AMPA receptor subunit
Abstract
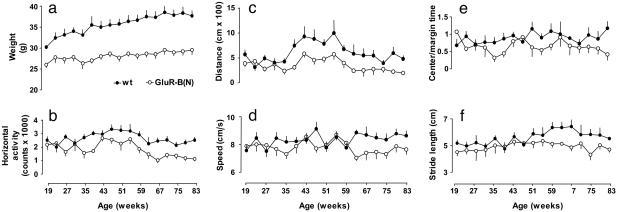
Amyotrophic lateral sclerosis (ALS) is a devastating disorder of the central nervous system in middle and old age that leads to progressive loss of spinal motoneurons. Transgenic mice overexpressing mutated human Cu(2+)/Zn(2+) superoxide dismutase 1 (SOD1) reproduce clinical features of the familial form of ALS. However, changes in SOD1 activity do not correlate with severity of motor decline in sporadic cases, indicating that targets unrelated to superoxide metabolism contribute to the pathogenesis of the disease. We show here that transgenic expression in mice of GluR-B(N)-containing L-alpha-amino-3-hydroxy-5-methylisoxazole-4-proprionate (AMPA) receptors with increased Ca(2+) permeability leads to late-onset degeneration of neurons in the spinal cord and decline of motor functions. Neuronal death progresses over the entire lifespan but manifests clinically in late adulthood, resembling the course of a slow neurodegenerative disorder. Additional transgenic expression of mutated human SOD1 accelerates disease progression, aggravates the severity of motor decline, and decreases survival. These observations link persistently elevated Ca(2+) influx through AMPA channels with progressive motor decline and late-onset degeneration of spinal motoneurons, indicating that functionally altered AMPA channels may be causally related to pathogenesis of sporadic ALS in humans.
Figures



References
-
- Rowland, L. P. & Schneider, N. A. (2001) N. Engl. J. Med. 344, 1688–1700. - PubMed
-
- Bruijn, L. I., Miller, T. M. & Cleveland, D. W. (2004) Annu. Rev. Neurosci. 27, 723–749. - PubMed
-
- Couratier, P., Hugon, J., Sindou, P., Vallat, J. M. & Dumas, M. (1993) Lancet 341, 265–268. - PubMed
-
- Kawahara, Y., Ito, K., Sun, H., Aizawa, H., Kanazawa, I. & Kwak, S. (2004) Nature 427, 801. - PubMed
-
- Burnashev, N., Monyer, H., Seeburg, P. H. & Sakmann, B. (1992) Neuron 8, 189–198. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
