

Rift valley fever virus nonstructural protein NSs promotes viral RNA replication and transcription in a minigenome system
- PMID: 15827175
- PMCID: PMC1082746
- DOI: 10.1128/JVI.79.9.5606-5615.2005
Rift valley fever virus nonstructural protein NSs promotes viral RNA replication and transcription in a minigenome system
Abstract
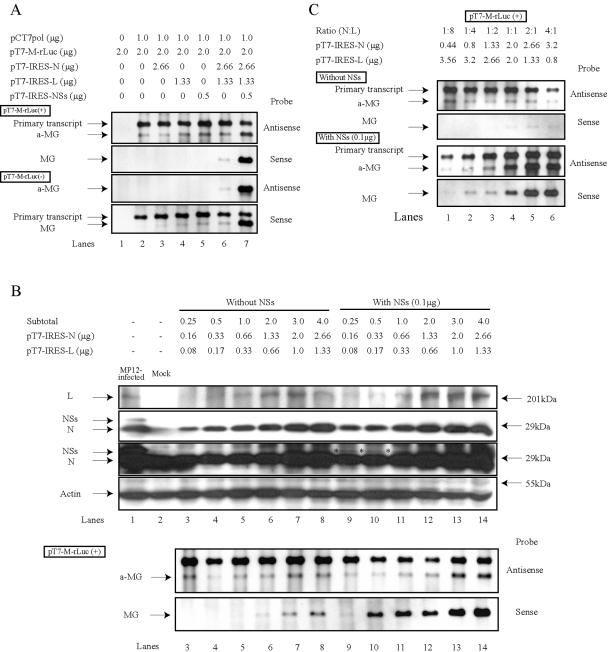
Rift Valley fever virus (RVFV), which belongs to the genus Phlebovirus, family Bunyaviridae, has a tripartite negative-strand genome (S, M, and L segments) and is an important mosquito-borne pathogen for domestic animals and humans. We established an RVFV T7 RNA polymerase-driven minigenome system in which T7 RNA polymerase from an expression plasmid drove expression of RNA transcripts for viral proteins and minigenome RNA transcripts carrying a reporter gene between both termini of the M RNA segment in 293T cells. Like other viruses of the Bunyaviridae family, replication and transcription of the RVFV minigenome required expression of viral N and L proteins. Unexpectedly, the coexpression of an RVFV nonstructural protein, NSs, with N and L proteins resulted in a significant enhancement of minigenome RNA replication. Coexpression of NSs protein with N and L proteins also enhanced minigenome mRNA transcription in the cells expressing viral-sense minigenome RNA transcripts. NSs protein expression increased the RNA replication of minigenomes that originated from S and L RNA segments. Enhancement of minigenome RNA synthesis by NSs protein occurred in cells lacking alpha/beta interferon (IFN-alpha/beta) genes, indicating that the effect of NSs protein on minigenome RNA replication was unrelated to a putative NSs protein-induced inhibition of IFN-alpha/beta production. Our finding that RVFV NSs protein augmented minigenome RNA synthesis was in sharp contrast to reports that Bunyamwera virus (genus Bunyavirus) NSs protein inhibits viral minigenome RNA synthesis, suggesting that RVFV NSs protein and Bunyamwera virus NSs protein have distinctly different biological roles in viral RNA synthesis.
Figures
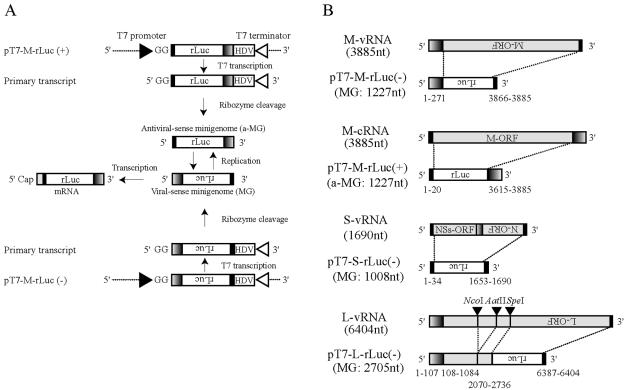
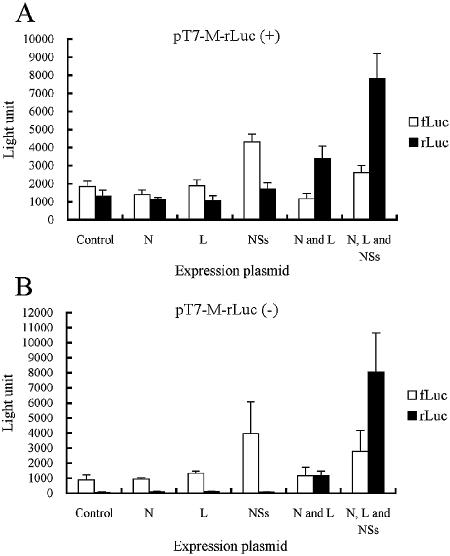


References
-
- Accardi, L., C. Prehaud, P. Di Bonito, S. Mochi, M. Bouloy, and C. Giorgi. 2001. Activity of Toscana and Rift Valley fever virus transcription complexes on heterologous templates. J. Gen. Virol. 82:781-785. - PubMed
-
- Balkhy, H. H., and Z. A. Memish. 2003. Rift Valley fever: an uninvited zoonosis in the Arabian peninsula. Int. J. Antimicrob. Agents 21:153-157. - PubMed
-
- Blakqori, G., G. Kochs, O. Haller, and F. Weber. 2003. Functional L polymerase of La Crosse virus allows in vivo reconstitution of recombinant nucleocapsids. J. Gen. Virol. 84:1207-1214. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
