

Arabidopsis CBF3/DREB1A and ABF3 in transgenic rice increased tolerance to abiotic stress without stunting growth
- PMID: 15834008
- PMCID: PMC1104188
- DOI: 10.1104/pp.104.059147
Arabidopsis CBF3/DREB1A and ABF3 in transgenic rice increased tolerance to abiotic stress without stunting growth
Abstract
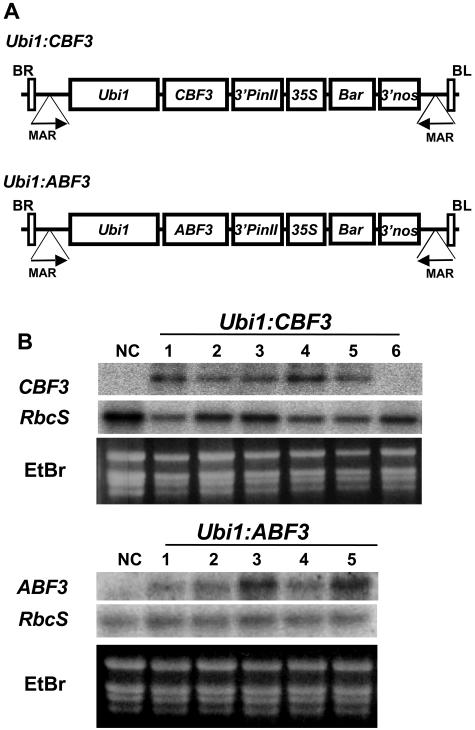
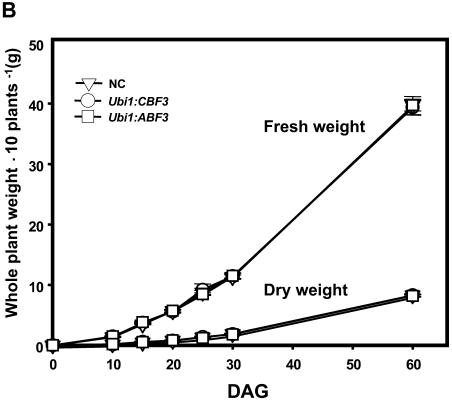
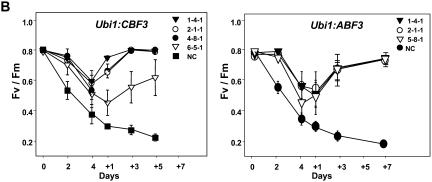
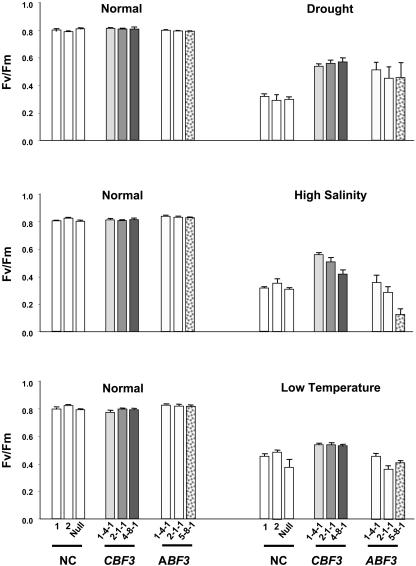
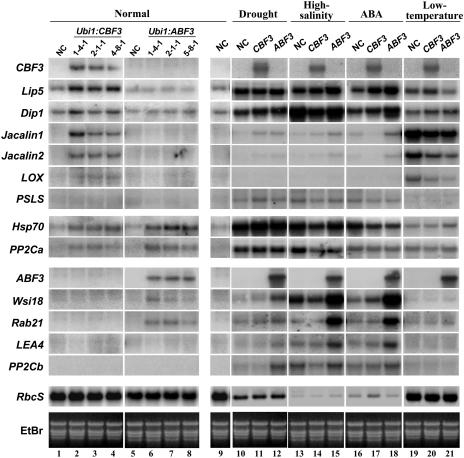
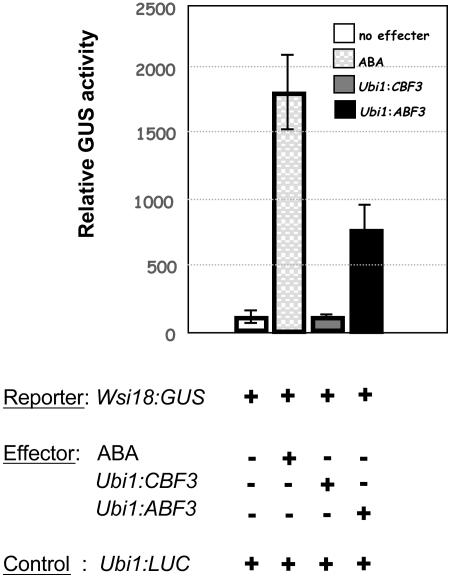
Rice (Oryza sativa), a monocotyledonous plant that does not cold acclimate, has evolved differently from Arabidopsis (Arabidopsis thaliana), which cold acclimates. To understand the stress response of rice in comparison with that of Arabidopsis, we developed transgenic rice plants that constitutively expressed CBF3/DREB1A (CBF3) and ABF3, Arabidopsis genes that function in abscisic acid-independent and abscisic acid-dependent stress-response pathways, respectively. CBF3 in transgenic rice elevated tolerance to drought and high salinity, and produced relatively low levels of tolerance to low-temperature exposure. These data were in direct contrast to CBF3 in Arabidopsis, which is known to function primarily to enhance freezing tolerance. ABF3 in transgenic rice increased tolerance to drought stress alone. By using the 60 K Rice Whole Genome Microarray and RNA gel-blot analyses, we identified 12 and 7 target genes that were activated in transgenic rice plants by CBF3 and ABF3, respectively, which appear to render the corresponding plants acclimated for stress conditions. The target genes together with 13 and 27 additional genes are induced further upon exposure to drought stress, consequently making the transgenic plants more tolerant to stress conditions. Interestingly, our transgenic plants exhibited neither growth inhibition nor visible phenotypic alterations despite constitutive expression of the CBF3 or ABF3, unlike the results previously obtained from Arabidopsis where transgenic plants were stunted.
Figures


References
-
- Baker SS, Wilhelm KS, Thomashow MF (1994) The 5′-region of Arabidopsis thaliana cor15a has cis-acting elements that confer cold-, drought- and ABA-regulated gene expression. Plant Mol Biol 24: 1–13 - PubMed
-
- Bray EA (1997) Plant responses to water deficit. Trends Plant Sci 2: 48–54
-
- Busk PK, Jensen AB, Pages M (1997) Regulatory elements in vivo in the promoter of the abscisic acid responsive gene rab17 from maize. Plant J 11: 1285–1295 - PubMed
-
- Choi HI, Hong JH, Ha JO, Kang JY, Kim SY (2000) ABFs, a family of ABA-responsive element binding factors. J Biol Chem 275: 1723–1730 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
