

Citrate secretion coupled with the modulation of soybean root tip under aluminum stress. Up-regulation of transcription, translation, and threonine-oriented phosphorylation of plasma membrane H+-ATPase
- PMID: 15834009
- PMCID: PMC1104183
- DOI: 10.1104/pp.104.058065
Citrate secretion coupled with the modulation of soybean root tip under aluminum stress. Up-regulation of transcription, translation, and threonine-oriented phosphorylation of plasma membrane H+-ATPase
Erratum in
- Plant Physiol. 2005 Sep;139(1):557. Hideo, Sasakawa [corrected to Sasakawa, Hideo]
Abstract
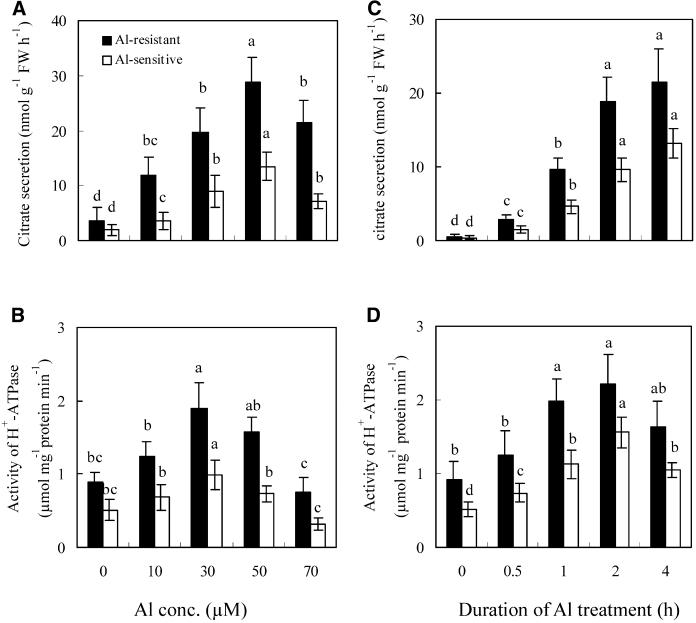
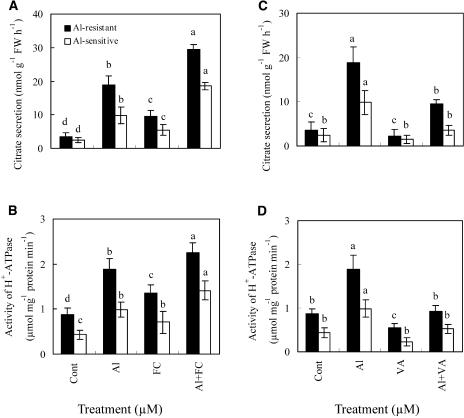
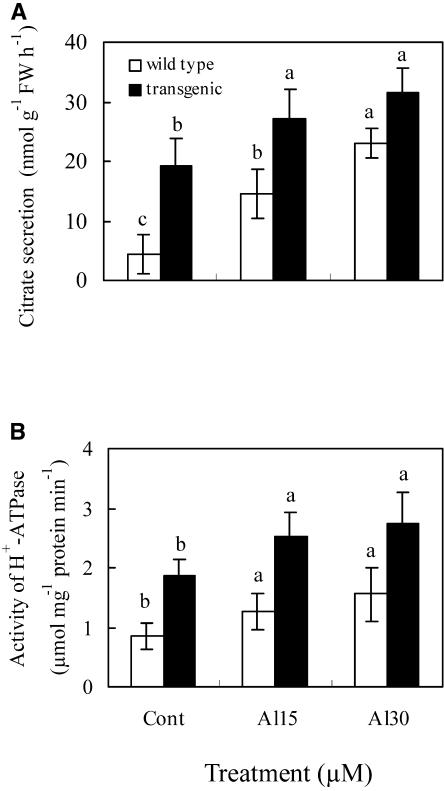
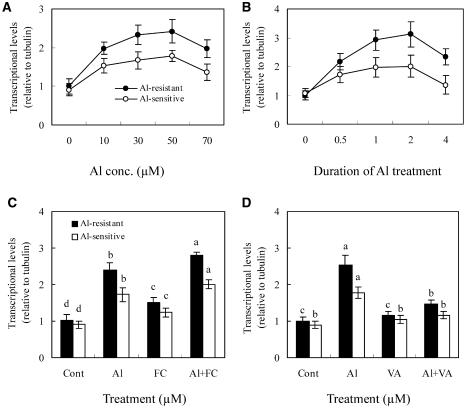
The aluminum (Al)-induced secretion of citrate has been regarded as an important mechanism for Al resistance in soybean (Glycine max). However, the mechanism of how Al induces citrate secretion remains unclear. In this study, we investigated the regulatory role of plasma membrane H+-ATPase on the Al-induced secretion of citrate from soybean roots. Experiments performed with plants grown in full nutrient solution showed that Al-induced activity of plasma membrane H+-ATPase paralleled secretion of citrate. Vanadate and fusicoccin, an inhibitor and an activator, respectively, of plasma membrane H+-ATPase, exerted inhibitory and stimulatory effects on the Al-induced secretion of citrate. Higher activity of plasma membrane H+-ATPase coincided with more citrate secretion in Al-resistant than Al-sensitive soybean cultivars. These results suggested that the effects of Al stress on citrate secretion were mediated via modulation of the activity of plasma membrane H+-ATPase. The relationship between the Al-induced secretion of citrate and the activity of plasma membrane H+-ATPase was further demonstrated by analysis of plasma membrane H+-ATPase transgenic Arabidopsis (Arabidopsis thaliana). When plants were grown on Murashige and Skoog medium containing 30 microM Al (9.1 microM Al3+ activity), transgenic plants exuded more citrate compared with wild-type Arabidopsis. Results from real-time reverse transcription-PCR and immunodetection analysis indicated that the increase of plasma membrane H+-ATPase activity by Al is caused by transcriptional and translational regulation. Furthermore, plasma membrane H+-ATPase activity and expression were higher in an Al-resistant cultivar than in an Al-sensitive cultivar. Al activated the threonine-oriented phosphorylation of plasma membrane H+-ATPase in a dose- and time-dependent manner. Taken together, our results demonstrated that up-regulation of plasma membrane H+-ATPase activity was associated with the secretion of citrate from soybean roots.
Figures
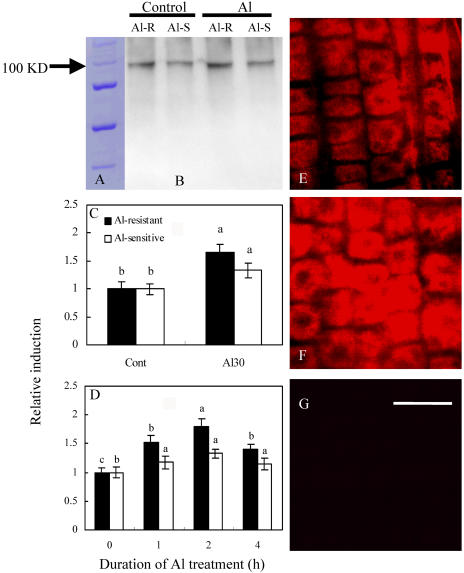
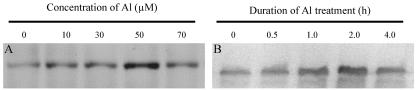
References
-
- Ahn SJ, Rengel Z, Matsumoto H (2004) Aluminum-induced plasma membrane surface potential and H+-ATPase activity in near-isogenic wheat lines differing in tolerance to aluminum. New Phytol 162: 71–79
-
- Arango M, Gevaudant F, Oufattole M, Boutry M (2003) The plasma membrane proton pump ATPase: the significance of gene subfamilies. Planta 216: 355–365 - PubMed
-
- Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248–254 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
