

Ethylene insensitivity conferred by the Green-ripe and Never-ripe 2 ripening mutants of tomato
- PMID: 15834010
- PMCID: PMC1104181
- DOI: 10.1104/pp.104.057745
Ethylene insensitivity conferred by the Green-ripe and Never-ripe 2 ripening mutants of tomato
Abstract
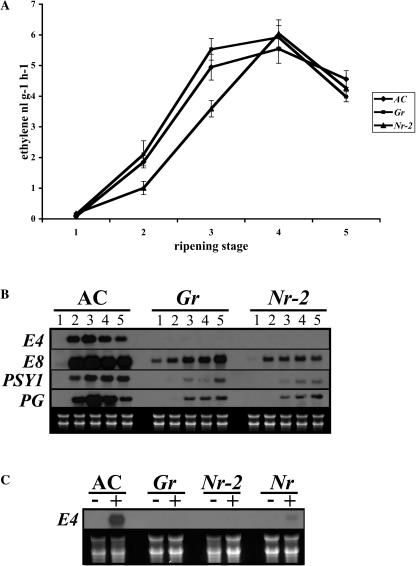
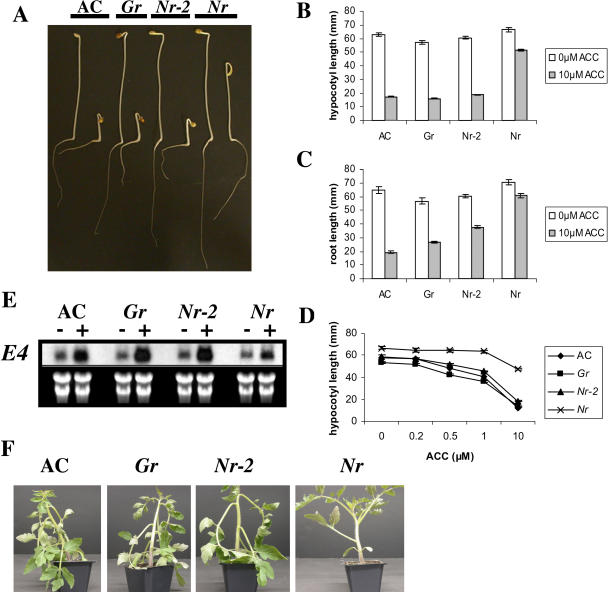
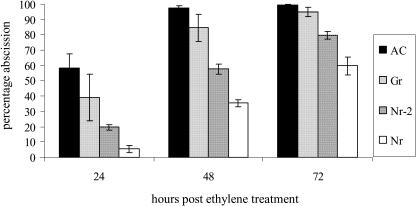
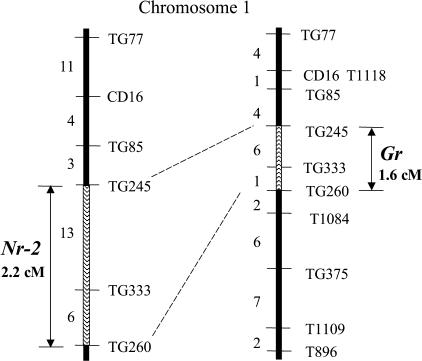
The ripening of a fleshy fruit represents the summation of an array of biochemical processes that are regulated by interactions between developmental programs and environmental inputs. Analysis of tomato (Solanum lycopersicum) mutants and inhibitor studies indicate that ethylene is necessary for full development of the ripening program of climacteric fruit such as tomato, yet ethylene alone is not sufficient. This suggests that an interaction between ethylene and nonethylene (or developmental) pathways mediates ripening. In this study, we have examined the physiological basis for ripening inhibition of the dominant Green-ripe (Gr) and Never-ripe 2 (Nr-2) mutants of tomato. Our data suggest that this inhibition is due to ethylene insensitivity in mutant fruit. Further investigation of ethylene responses in Gr and Nr-2 plants also revealed weak ethylene insensitivity during floral senescence and abscission and, during inhibition of root elongation, a phenotype associated with the triple response. However, ethylene-induced inhibition of hypocotyl elongation and petiole epinasty are normal in Gr and Nr-2, suggesting that these loci regulate a subset of ethylene responses. We have mapped both dominant mutations to a 2-cM overlapping region of the long arm of chromosome 1 of tomato, a region not previously linked to any known ethylene signaling loci. The phenotypic similarity and overlapping map location of these mutations suggest Gr and Nr-2 may be allelic and may possibly encode a novel component of the ethylene response pathway.
Figures
References
-
- Adams-Phillips L, Barry C, Giovannoni J (2004. a) Signal transduction systems regulating fruit ripening. Trends Plant Sci 9: 331–338 - PubMed
-
- Adams-Phillips L, Barry C, Kannan P, Leclercq J, Bouzayen M, Giovannoni J (2004. b) Evidence that CTR1-mediated ethylene signal transduction in tomato is encoded by a multigene family whose members display distinct regulatory features. Plant Mol Biol 54: 387–404 - PubMed
-
- Darby LA, Ritchie DB, Taylor IB (1977) Isogenic lines of the tomato “Ailsa Craig”. Annual Report Glasshouse Crops Research Institute, Littlehampton, UK, pp 168–184
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
