

IL-7 receptor signaling is necessary for stage transition in adult B cell development through up-regulation of EBF
- PMID: 15837809
- PMCID: PMC2213146
- DOI: 10.1084/jem.20050158
IL-7 receptor signaling is necessary for stage transition in adult B cell development through up-regulation of EBF
Abstract
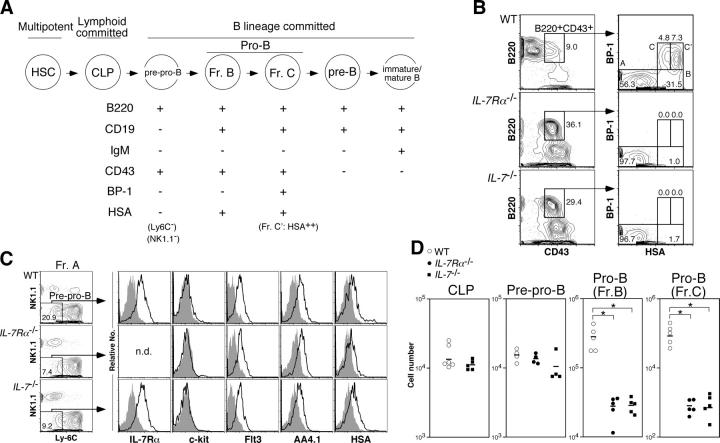
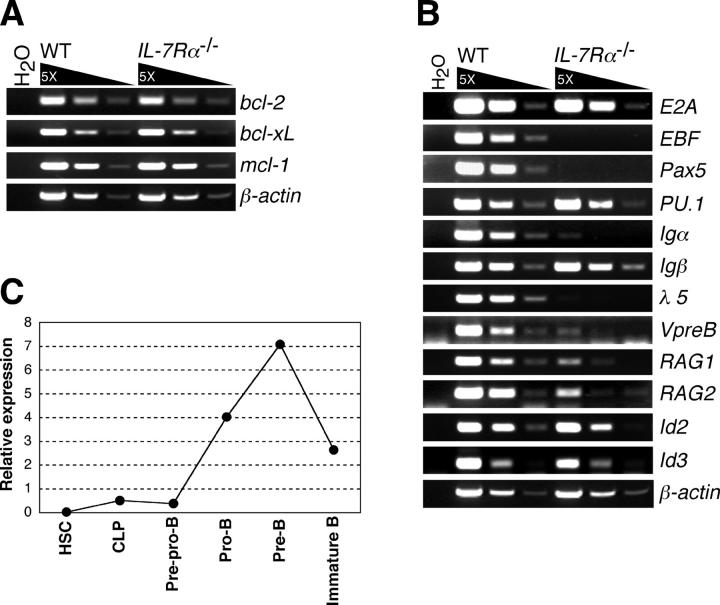
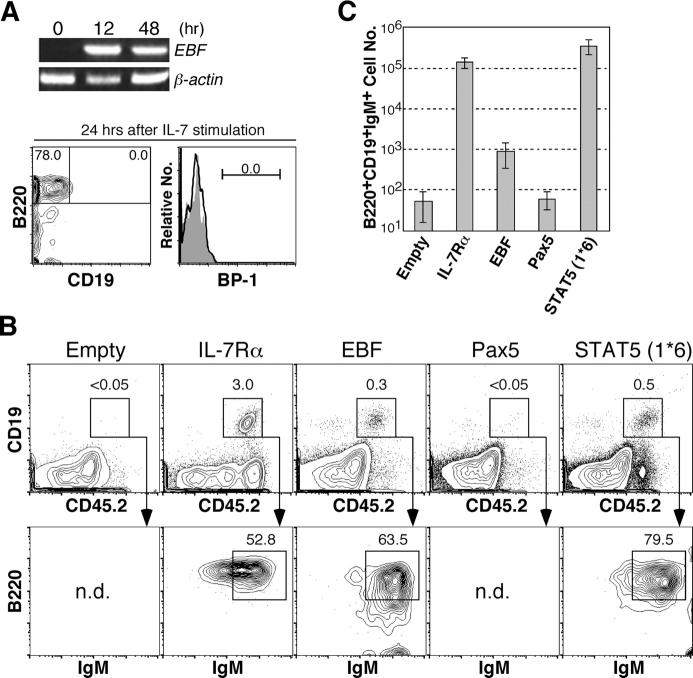
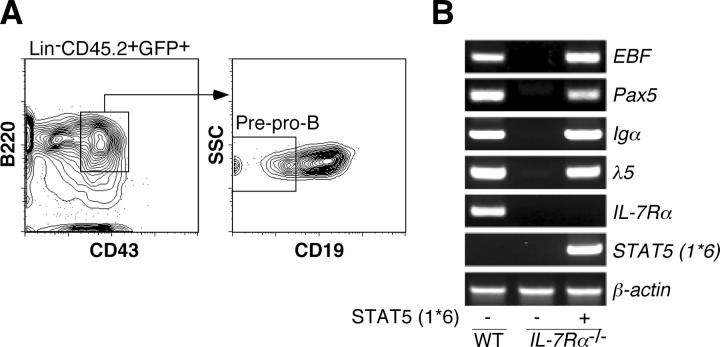
Cytokine receptor signals have been suggested to stimulate cell differentiation during hemato/lymphopoiesis. Such action, however, has not been clearly demonstrated. Here, we show that adult B cell development in IL-7(-/-) and IL-7R alpha(2/-) mice is arrested at the pre-pro-B cell stage due to insufficient expression of the B cell-specific transcription factor EBF and its target genes, which form a transcription factor network in determining B lineage specification. EBF expression is restored in IL-7(-/-) pre-pro-B cells upon IL-7 stimulation or in IL-7R alpha(-/-) pre-pro-B cells by activation of STAT5, a major signaling molecule downstream of the IL-7R signaling pathway. Furthermore, enforced EBF expression partially rescues B cell development in IL-7R alpha(-/-) mice. Thus, IL-7 receptor signaling is a participant in the formation of the transcription factor network during B lymphopoiesis by up-regulating EBF, allowing stage transition from the pre-pro-B to further maturational stages.
Figures
References
-
- Quong, M.W., W.J. Romanow, and C. Murre. 2002. E protein function in lymphocyte development. Annu. Rev. Immunol. 20:301–322. - PubMed
-
- Busslinger, M. 2004. Transcriptional control of early B cell development. Annu. Rev. Immunol. 22:55–79. - PubMed
-
- Medina, K.L., J.M. Pongubala, K.L. Reddy, D.W. Lancki, R. Dekoter, M. Kieslinger, R. Grosschedl, and H. Singh. 2004. Assembling a gene regulatory network for specification of the B cell fate. Dev. Cell. 7:607–617. - PubMed
-
- O'Riordan, M., and R. Grosschedl. 1999. Coordinate regulation of B cell differentiation by the transcription factors EBF and E2A. Immunity. 11:21–31. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
