

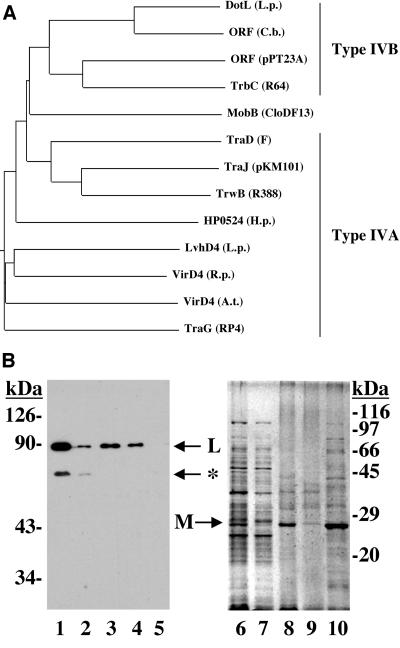
The DotL protein, a member of the TraG-coupling protein family, is essential for Viability of Legionella pneumophila strain Lp02
- PMID: 15838018
- PMCID: PMC1082803
- DOI: 10.1128/JB.187.9.2927-2938.2005
The DotL protein, a member of the TraG-coupling protein family, is essential for Viability of Legionella pneumophila strain Lp02
Abstract
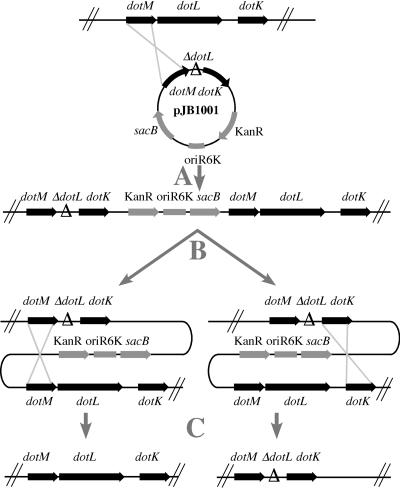
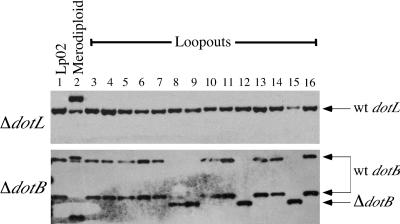
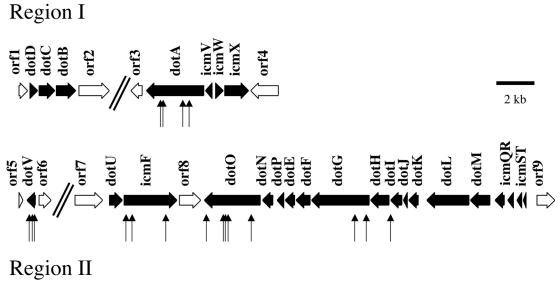
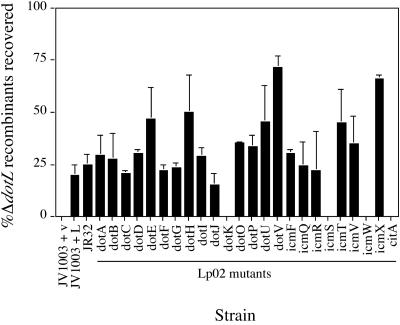
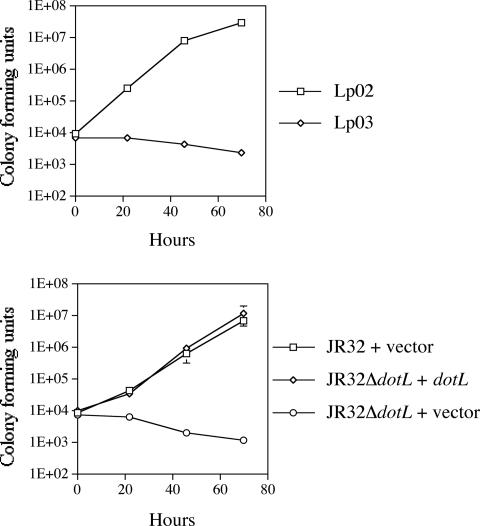
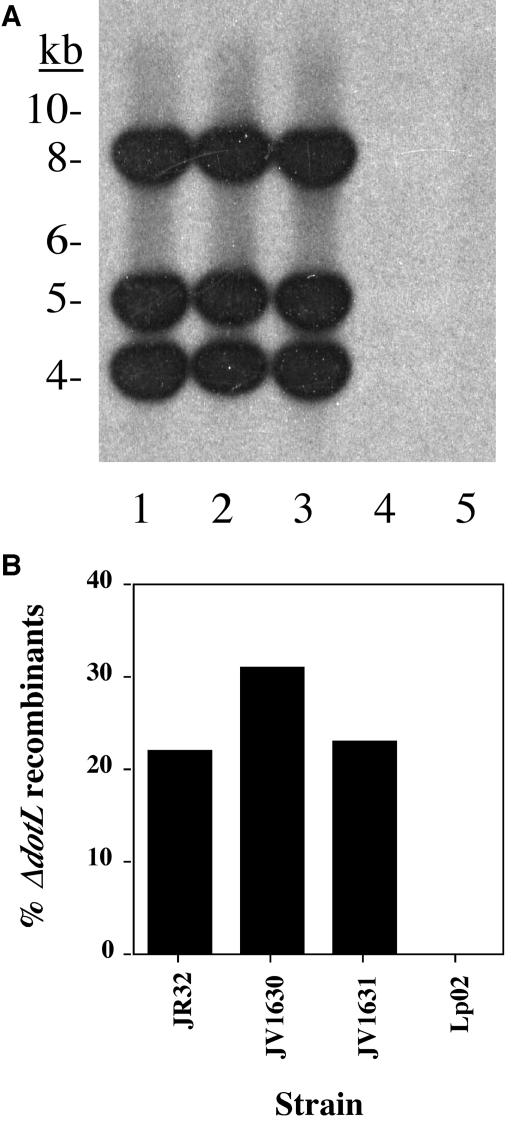
Legionella pneumophila is able to survive inside phagocytic cells by an internalization route that bypasses fusion of the nascent phagosome with the endocytic pathway to allow formation of a replicative phagosome. The dot/icm genes, a major virulence system of L. pneumophila, encode a type IVB secretion system that is required for intracellular growth. One Dot protein, DotL, has sequence similarity to type IV secretion system coupling proteins (T4CPs). In other systems, coupling proteins are not required for viability of the organism. Here we report the first example of a strain, L. pneumophila Lp02, in which a putative T4CP is essential for viability of the organism on bacteriological media. This result is particularly surprising since the majority of the dot/icm genes in Lp02 are dispensable for growth outside of a host cell, a condition that does not require a functional Dot/Icm secretion complex. We were able to isolate suppressors of the Delta dotL lethality and found that many contained mutations in other components of the Dot/Icm secretion system. A systematic analysis of dot/icm deletion mutants revealed that the majority of them (20 of 26) suppressed the lethality phenotype, indicating a partially assembled secretion system may be the source of Delta dotL toxicity in the wild-type strain. These results are consistent with a model in which the DotL protein plays a role in regulating the activity of the L. pneumophila type IV secretion apparatus.
Figures


References
-
- Berger, K. H., and R. R. Isberg. 1993. Two distinct defects in intracellular growth complemented by a single genetic locus in Legionella pneumophila. Mol. Microbiol. 7:7-19. - PubMed
-
- Blomfield, I. C., V. Vaughn, R. F. Rest, and B. I. Eisenstein. 1991. Allelic exchange in Escherichia coli using the Bacillus subtilis sacB gene and a temperature-sensitive pSC101 replicon. Mol. Microbiol. 5:1447-1457. - PubMed
-
- Brand, B. C., A. B. Sadosky, and H. A. Shuman. 1994. The Legionella pneumophila icm locus: a set of genes required for intracellular multiplication in human macrophages. Mol. Microbiol. 14:797-808. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
