

Role of HtrA in growth and competence of Streptococcus mutans UA159
- PMID: 15838029
- PMCID: PMC1082816
- DOI: 10.1128/JB.187.9.3028-3038.2005
Role of HtrA in growth and competence of Streptococcus mutans UA159
Abstract
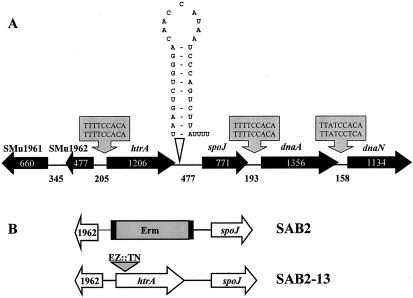
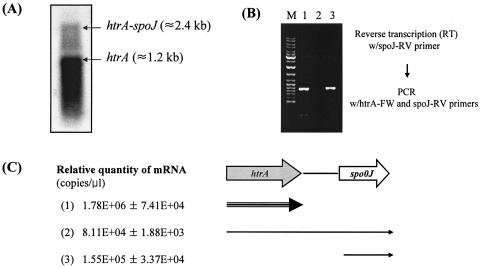
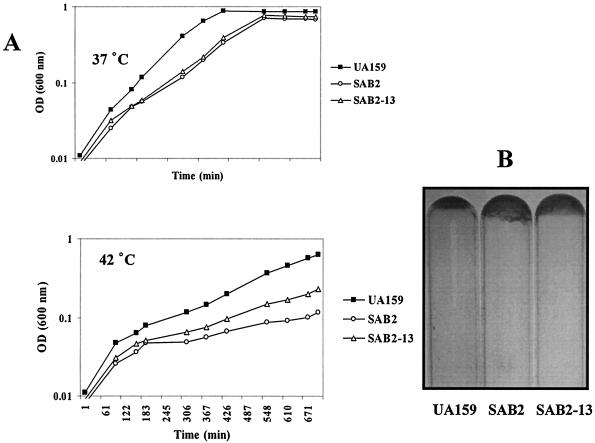
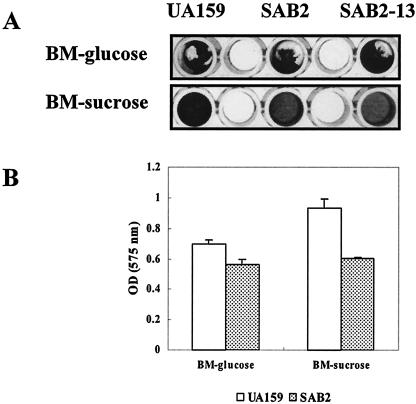
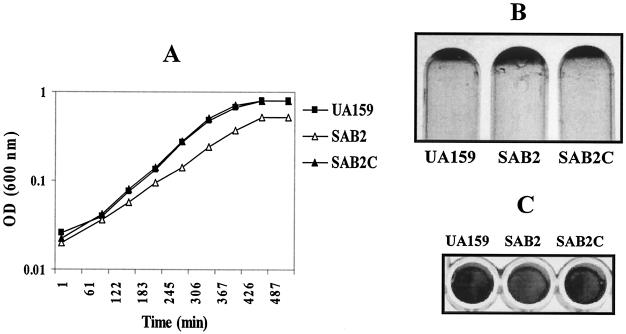
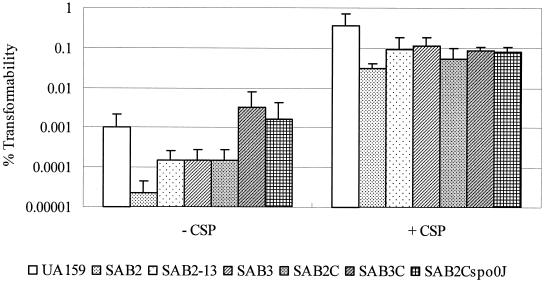
We report here that HtrA plays a role in controlling growth and competence development for genetic transformation in Streptococcus mutans. Disruption of the gene for HtrA resulted in slow growth at 37 degrees C, reduced thermal tolerance at 42 degrees C, and altered sucrose-dependent biofilm formation on polystyrene surfaces. The htrA mutant also displayed a significantly reduced ability to undergo genetic transformation. A direct association between HtrA and genetic competence was demonstrated by the increased expression of the htrA gene upon exposure to competence-stimulating peptide. The induction of htrA gradually reached a maximum at around 20 min, suggesting that HtrA may be involved in a late competence response. Complementation of the htrA mutation in a single copy on the chromosome of the mutant could rescue the defective growth phenotypes but not transformability, apparently because a second gene, spo0J, immediately downstream of htrA, also affects transformation. The htrA and spo0J genes were shown to be both individually transcribed and cotranscribed and probably have a functional connection in competence development. HtrA regulation appears to be finely tuned in S. mutans, since strains containing multiple copies of htrA exhibited abnormal growth phenotypes. Collectively, the results reveal HtrA to be an integral component of the regulatory network connecting cellular growth, stress tolerance, biofilm formation, and competence development and reveal a novel role for the spo0J gene in genetic transformation.
Figures
References
-
- Ajdic, D., W. M. McShan, R. E. McLaughlin, G. Savic, J. Chang, M. B. Carson, C. Primeaux, R. Tian, S. Kenton, H. Jia, S. Lin, Y. Qian, S. Li, H. Zhu, F. Najar, H. Lai, J. White, B. A. Roe, and J. J. Ferretti. 2002. Genome sequence of Streptococcus mutans UA159, a cariogenic dental pathogen. Proc. Natl. Acad. Sci. USA 99:14434-14439. - PMC - PubMed
-
- Allaker, R. P., S. V. Seddon, C. Tredwin, and E. Lynch. 1998. Detection of Streptococcus mutans by PCR amplification of the spaP gene in teeth rendered caries free. J. Dent. 26:443-445. - PubMed
-
- Autret, S., R. Nair, and J. Errington. 2001. Genetic analysis of the chromosome segregation protein Spo0J of Bacillus subtilis: evidence for separate domains involved in DNA binding and interactions with Soj protein. Mol. Microbiol. 41:743-755. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
