

Synthesis of autoinducer 2 by the lyme disease spirochete, Borrelia burgdorferi
- PMID: 15838035
- PMCID: PMC1082833
- DOI: 10.1128/JB.187.9.3079-3087.2005
Synthesis of autoinducer 2 by the lyme disease spirochete, Borrelia burgdorferi
Abstract
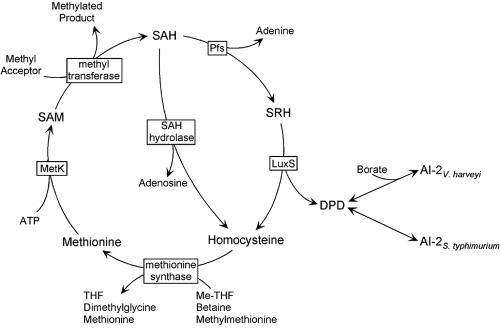
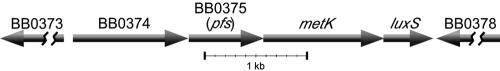
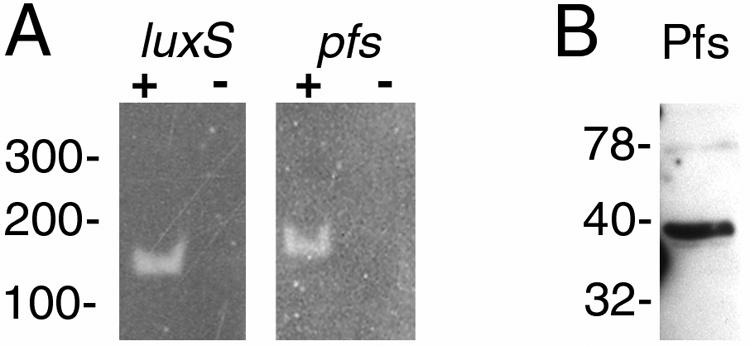
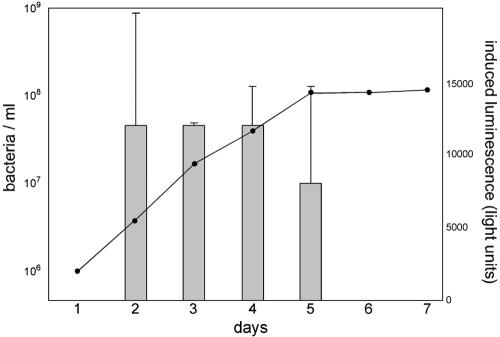
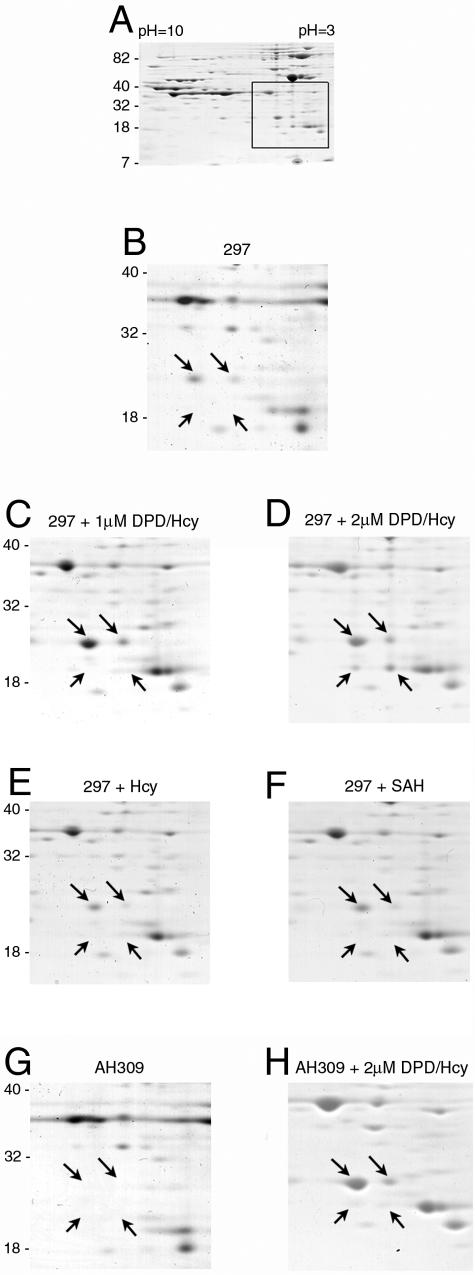
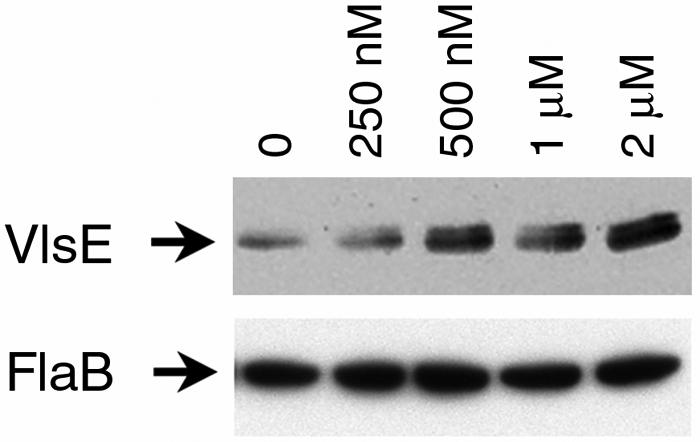
Defining the metabolic capabilities and regulatory mechanisms controlling gene expression is a valuable step in understanding the pathogenic properties of infectious agents such as Borrelia burgdorferi. The present studies demonstrated that B. burgdorferi encodes functional Pfs and LuxS enzymes for the breakdown of toxic products of methylation reactions. Consistent with those observations, B. burgdorferi was shown to synthesize the end product 4,5-dihydroxy-2,3-pentanedione (DPD) during laboratory cultivation. DPD undergoes spontaneous rearrangements to produce a class of pheromones collectively named autoinducer 2 (AI-2). Addition of in vitro-synthesized DPD to cultured B. burgdorferi resulted in differential expression of a distinct subset of proteins, including the outer surface lipoprotein VlsE. Although many bacteria can utilize the other LuxS product, homocysteine, for regeneration of methionine, B. burgdorferi was found to lack such ability. It is hypothesized that B. burgdorferi produces LuxS for the express purpose of synthesizing DPD and utilizes a form of that molecule as an AI-2 pheromone to control gene expression.
Figures
References
-
- Anguita, J., M. N. Hedrick, and E. Fikrig. 2003. Adaptation of Borrelia burgdorferi in the tick and the mammalian host. FEMS Microbiol. Rev. 27:493-504. - PubMed
-
- Bassler, B. L., M. Wright, and M. R. Silverman. 1994. Multiple signalling systems controlling expression of luminescence in Vibrio harveyi: sequence and function of genes encoding a second sensory pathway. Mol. Microbiol. 13:273-286. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
