

Plasmid-encoded regulator of extracellular proteases in Bacillus anthracis
- PMID: 15838040
- PMCID: PMC1082805
- DOI: 10.1128/JB.187.9.3133-3138.2005
Plasmid-encoded regulator of extracellular proteases in Bacillus anthracis
Abstract
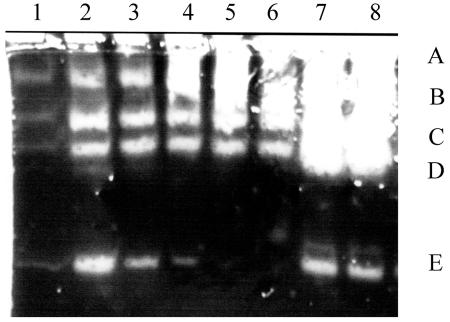
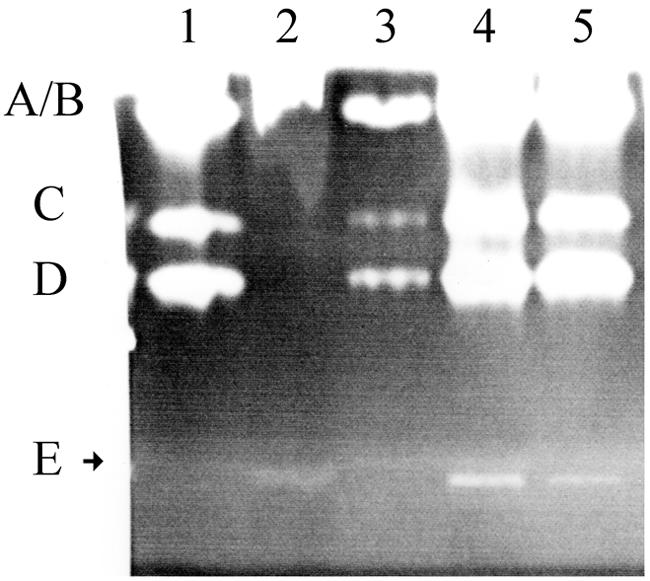
Bacillus anthracis Sterne cured of the pXO1 plasmid had enhanced secreted protease activity during the postexponential phase but no change in hemolytic or lecithinase activities. A zymogen profile revealed at least six proteases, including serine, metal, and perhaps cysteine types. There were similar amounts of protease secreted by the closely related species Bacillus cereus and Bacillus thuringiensis, but the patterns differed. Among the pXO1 plasmid-encoded proteins, there is a tetratricopeptide protein designated Cot43 that is related to the Rap proteins of Bacillus subtilis and the PlcR pleiotropic regulator of secreted enzymes and toxins in B. thuringiensis. A disruption of the cot43 gene resulted in overproduction of several proteases to a somewhat greater extent than in the plasmid-cured strain. Transformation of either of these strains with a clone of the cot43 gene resulted in the inhibition of accumulation of some of the proteases and induction of at least one. On the basis of lacZ fusions, transcription of the cot43 gene increased in late exponential cells at the time of protease accumulation. The expression of lacZ fusions to the upstream regions of two B. anthracis extracellular protease genes was greater in the strain with the disruption of cot43 than in the Sterne strain, indicating regulation at the level of transcription. In B. anthracis, a pXO1 plasmid-encoded protein directly modulates or indirectly regulates the transcription of genes for several chromosomally encoded extracellular proteases.
Figures
References
-
- Agaisse, H., and D. Lereclus. 1994. Structural and functional analysis of the promoter region involved in full expression of the cryIIIA toxin gene of Bacillus thuringiensis. Mol. Microbiol. 13:97-107. - PubMed
-
- Agaisse H., M. Gominet, O. A. Okstad, A.-B. Kolsto, and D. Lereclus. 1999. PlcR is a pleiotropic regulator of extracellular virulence factor gene expression in Bacillus thuringiensis. Mol. Microbiol. 32:1043-1053. - PubMed
-
- Arantes, O., and D. Lereclus. 1991. Construction of cloning vectors for Bacillus thuringiensis. Gene 108:115-119. - PubMed
-
- Bendtsen, J. D., H. Nielsen, G. von Heinje, and S. Brunek. 2004. Improved prediction of signal peptides: signal IP30. J. Mol. Biol. 340:783-795. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
