

Thrombin-induced oxidative stress contributes to the death of hippocampal neurons in vivo: role of microglial NADPH oxidase
- PMID: 15843610
- PMCID: PMC6724962
- DOI: 10.1523/JNEUROSCI.4306-04.2005
Thrombin-induced oxidative stress contributes to the death of hippocampal neurons in vivo: role of microglial NADPH oxidase
Abstract
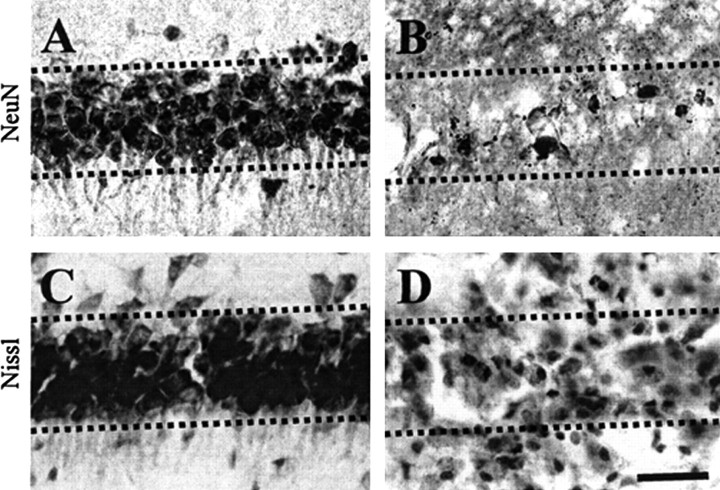
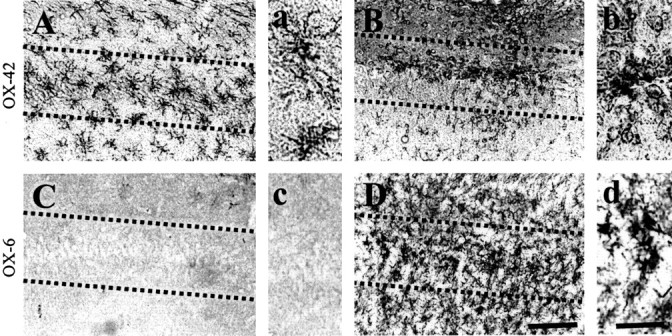
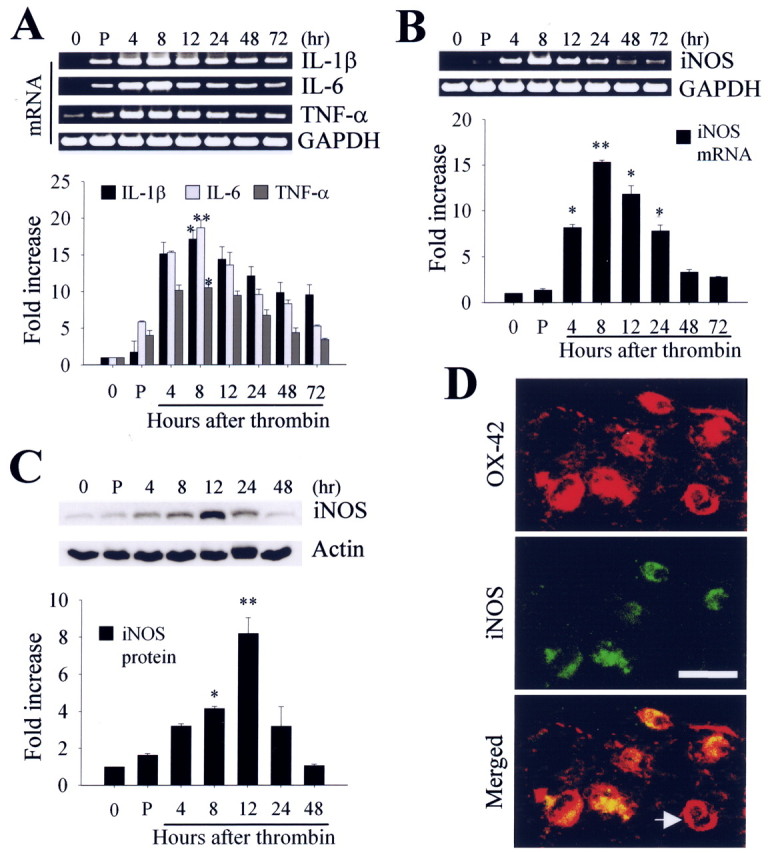
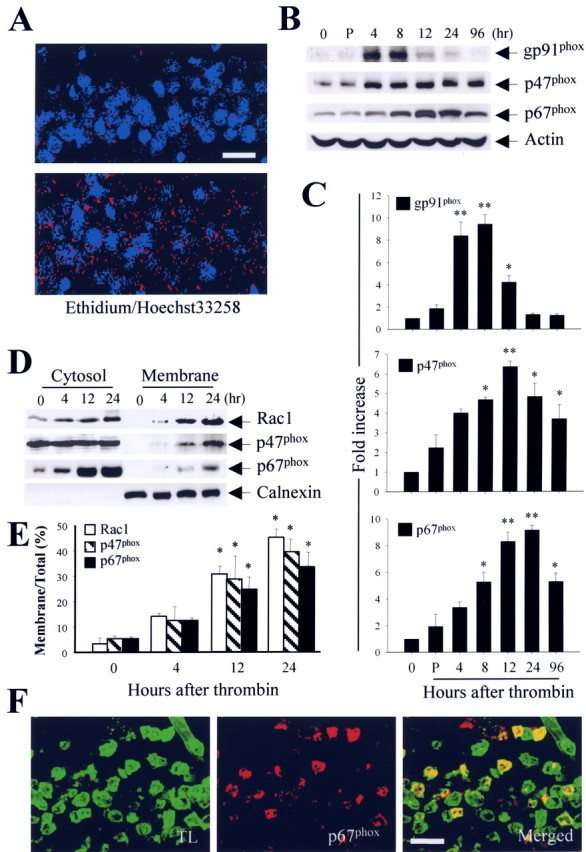
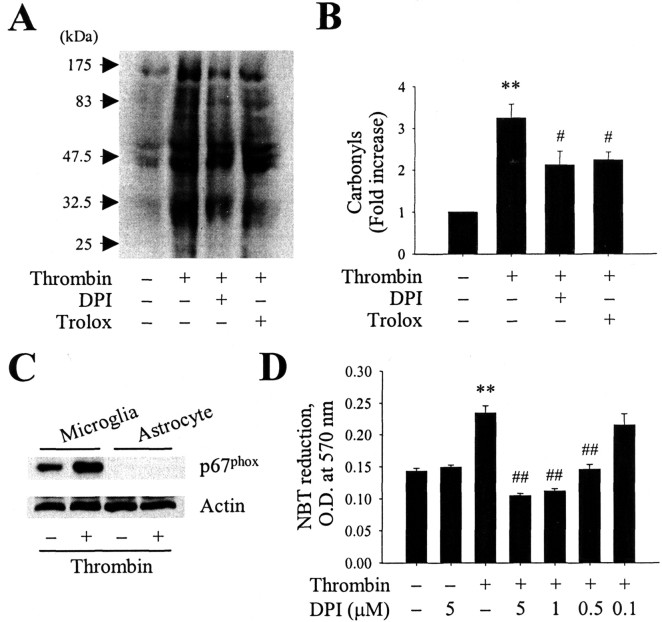
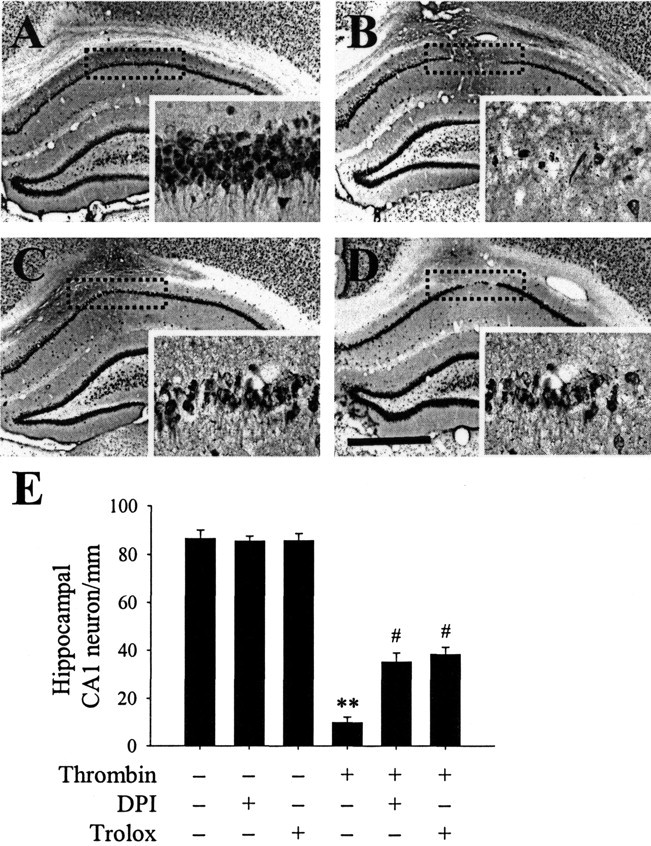
The present study investigated whether thrombin, a potent microglial activator, can induce reactive oxygen species (ROS) generation through activation of microglial NADPH oxidase and if this may contribute to oxidative damage and consequent neurodegeneration. Seven days after intrahippocampal injection of thrombin, Nissl staining and immunohistochemistry using the neuronal-specific nuclear protein NeuN revealed a significant loss in hippocampal CA1 neurons. In parallel, thrombin-activated microglia, assessed by OX-42 and OX-6 immunohistochemistry, and ROS production, assessed by hydroethidine histochemistry, were observed in the hippocampal CA1 area in which degeneration of hippocampal neurons occurred. Reverse transcription-PCR at various time points after thrombin administration demonstrated an early and transient expression of inducible nitric oxide synthase (iNOS) and several proinflammatory cytokines. Western blot analysis and double-label immunohistochemistry showed an increase in the expression of and the localization of iNOS within microglia. Additional studies demonstrated that thrombin induced the upregulation of membrane (gp91(phox)) and cytosolic (p47(phox) and p67(phox)) components, translocation of cytosolic proteins (p47(phox), p67(phox), and Rac1) to the membrane, and p67(phox) expression of the NADPH oxidase in microglia in the hippocampus in vivo, indicating the activation of NADPH oxidase. The thrombin-induced oxidation of proteins and loss of hippocampal CA1 neurons were partially inhibited by an NADPH oxidase inhibitor and by an antioxidant. To our knowledge, the present study is the first to demonstrate that thrombin-induced neurotoxicity in the hippocampus in vivo is caused by microglial NADPH oxidase-mediated oxidative stress. This suggests that thrombin inhibition or enhancing antioxidants may be beneficial for the treatment of neurodegenerative diseases, such as Alzheimer's disease, that are associated with microglial-derived oxidative damage.
Figures
Similar articles
-
Prothrombin kringle-2-induced oxidative stress contributes to the death of cortical neurons in vivo and in vitro: role of microglial NADPH oxidase.J Neuroimmunol. 2009 Sep 29;214(1-2):83-92. doi: 10.1016/j.jneuroim.2009.07.005. Epub 2009 Aug 5. J Neuroimmunol. 2009. PMID: 19660816
-
Thrombin-induced oxidative stress contributes to the death of hippocampal neurons: role of neuronal NADPH oxidase.J Neurosci Res. 2008 Apr;86(5):1053-63. doi: 10.1002/jnr.21571. J Neurosci Res. 2008. PMID: 18183616
-
Interleukin-4-induced oxidative stress via microglial NADPH oxidase contributes to the death of hippocampal neurons in vivo.Curr Aging Sci. 2008 Dec;1(3):192-201. doi: 10.2174/1874609810801030192. Curr Aging Sci. 2008. PMID: 20021392
-
Inflammatory neurodegeneration and mechanisms of microglial killing of neurons.Mol Neurobiol. 2010 Jun;41(2-3):242-7. doi: 10.1007/s12035-010-8105-9. Epub 2010 Mar 2. Mol Neurobiol. 2010. PMID: 20195798 Review.
-
NADPH oxidase as a therapeutic target in Alzheimer's disease.BMC Neurosci. 2008 Dec 3;9 Suppl 2(Suppl 2):S8. doi: 10.1186/1471-2202-9-S2-S8. BMC Neurosci. 2008. PMID: 19090996 Free PMC article. Review.
Cited by
-
Reactive oxygen species up-regulate CD11b in microglia via nitric oxide: Implications for neurodegenerative diseases.Free Radic Biol Med. 2008 Sep 1;45(5):686-99. doi: 10.1016/j.freeradbiomed.2008.05.026. Epub 2008 Jun 6. Free Radic Biol Med. 2008. PMID: 18590811 Free PMC article.
-
Monoamine Oxidase: A Potential Link in Papez Circuit to Generalized Anxiety Disorders.CNS Neurol Disord Drug Targets. 2024;23(5):638-655. doi: 10.2174/1871527322666230412105711. CNS Neurol Disord Drug Targets. 2024. PMID: 37055898 Review.
-
PPARalpha ligands inhibit radiation-induced microglial inflammatory responses by negatively regulating NF-kappaB and AP-1 pathways.Free Radic Biol Med. 2008 Dec 15;45(12):1695-704. doi: 10.1016/j.freeradbiomed.2008.09.002. Epub 2008 Sep 17. Free Radic Biol Med. 2008. PMID: 18852043 Free PMC article.
-
Features of microglia and neuroinflammation relevant to environmental exposure and neurotoxicity.Int J Environ Res Public Health. 2011 Jul;8(7):2980-3018. doi: 10.3390/ijerph8072980. Epub 2011 Jul 20. Int J Environ Res Public Health. 2011. PMID: 21845170 Free PMC article. Review.
-
Thrombin induces morphological and inflammatory astrocytic responses via activation of PAR1 receptor.Cell Death Discov. 2022 Apr 11;8(1):189. doi: 10.1038/s41420-022-00997-4. Cell Death Discov. 2022. PMID: 35399122 Free PMC article.
References
-
- Akiyama H, Ikeda K, Kondo H, McGeer PL (1992) Thrombin accumulation in brains of patients with Alzheimer's disease. Neurosci Lett 146: 152-154. - PubMed
-
- Akiyama H, Barger S, Barnum S, Bradt B, Bauer J, Cole GM, Cooper NR, Eikelenboom P, Emmerling M, Fiebich BL, Finch CE, Frautschy S, Griffin WS, Hampel H, Hull M, Landreth G, Lue L, Mrak R, Mackenzie IR, McGeer PL, et al. (2000) Inflammation and Alzheimer's disease. Neurobiol Aging 21: 383-421. - PMC - PubMed
-
- Babior BM (1999) NADPH oxidase: an update. Blood 93: 1464-1476. - PubMed
-
- Bal-Price A, Matthias A, Browin GC (2002) Stimulation of NADPH oxidase in activated rat microglia removes nitric oxide but induces peroxynitrite production. J Neurochem 80: 73-80. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous