

Requirement of Cul3 for axonal arborization and dendritic elaboration in Drosophila mushroom body neurons
- PMID: 15843622
- PMCID: PMC6724961
- DOI: 10.1523/JNEUROSCI.0149-05.2005
Requirement of Cul3 for axonal arborization and dendritic elaboration in Drosophila mushroom body neurons
Abstract
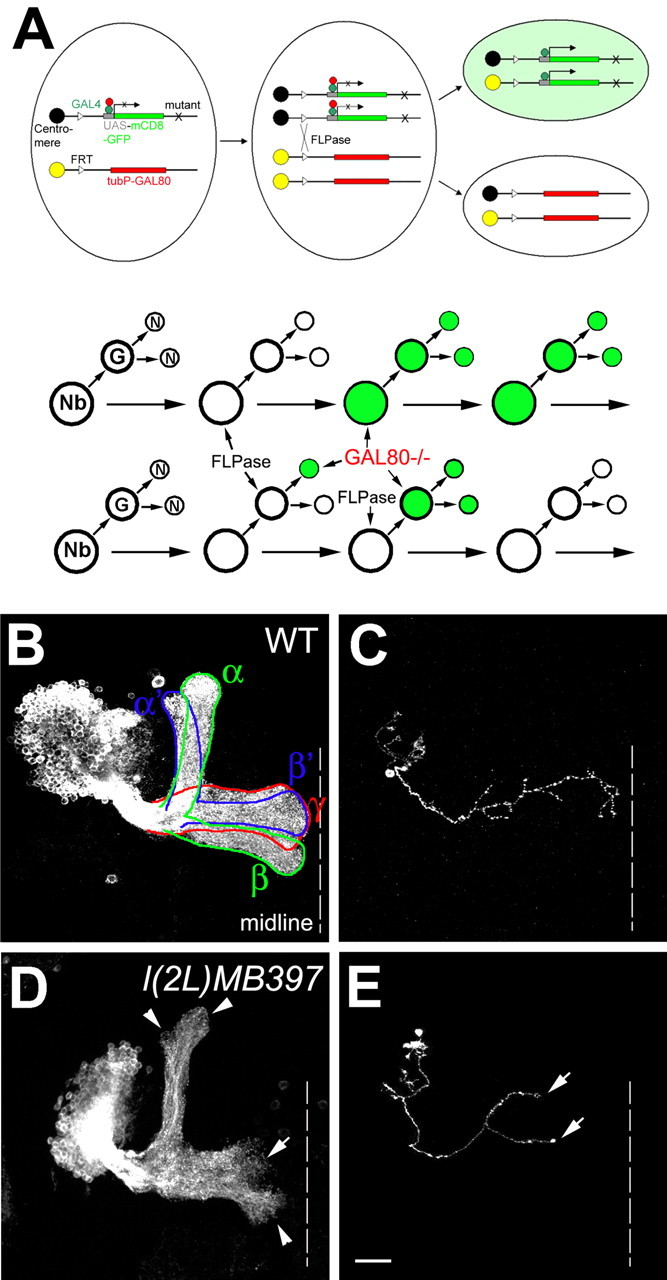
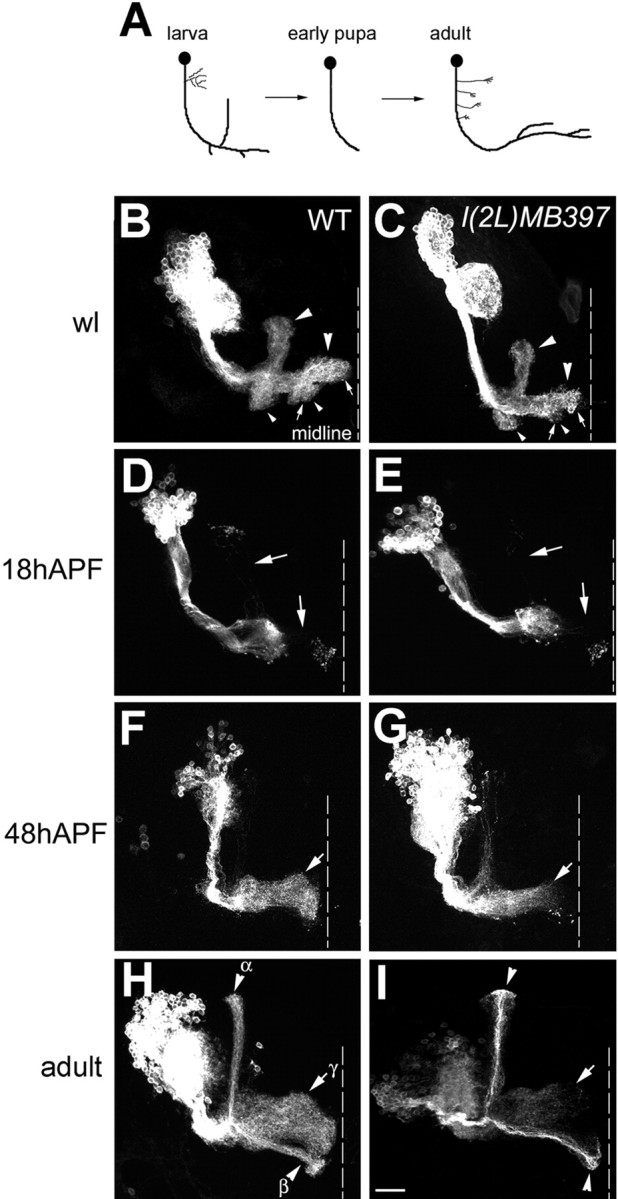
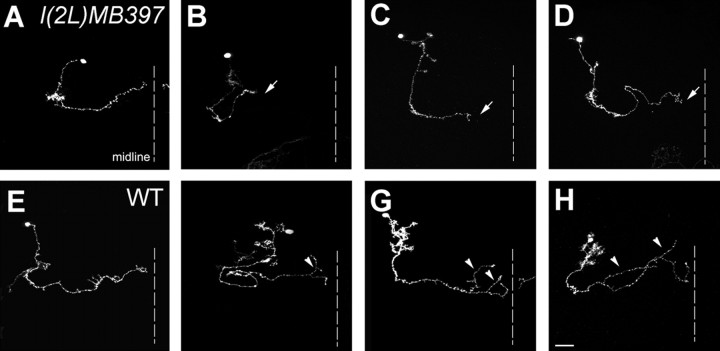
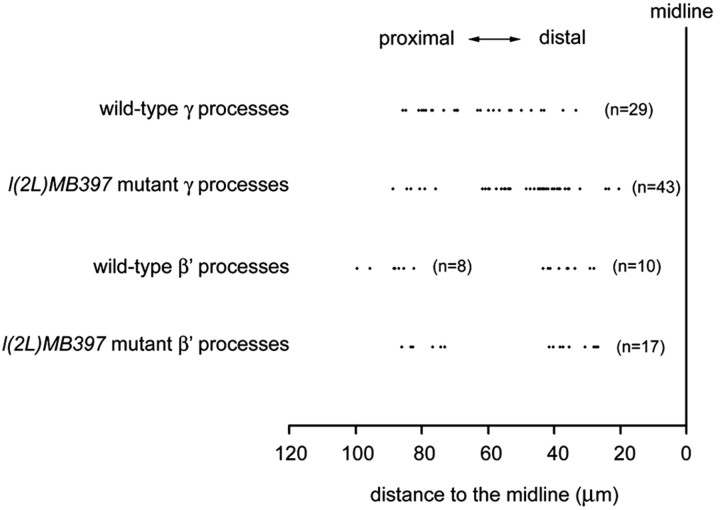
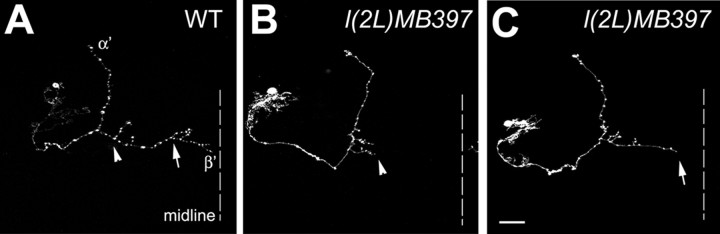
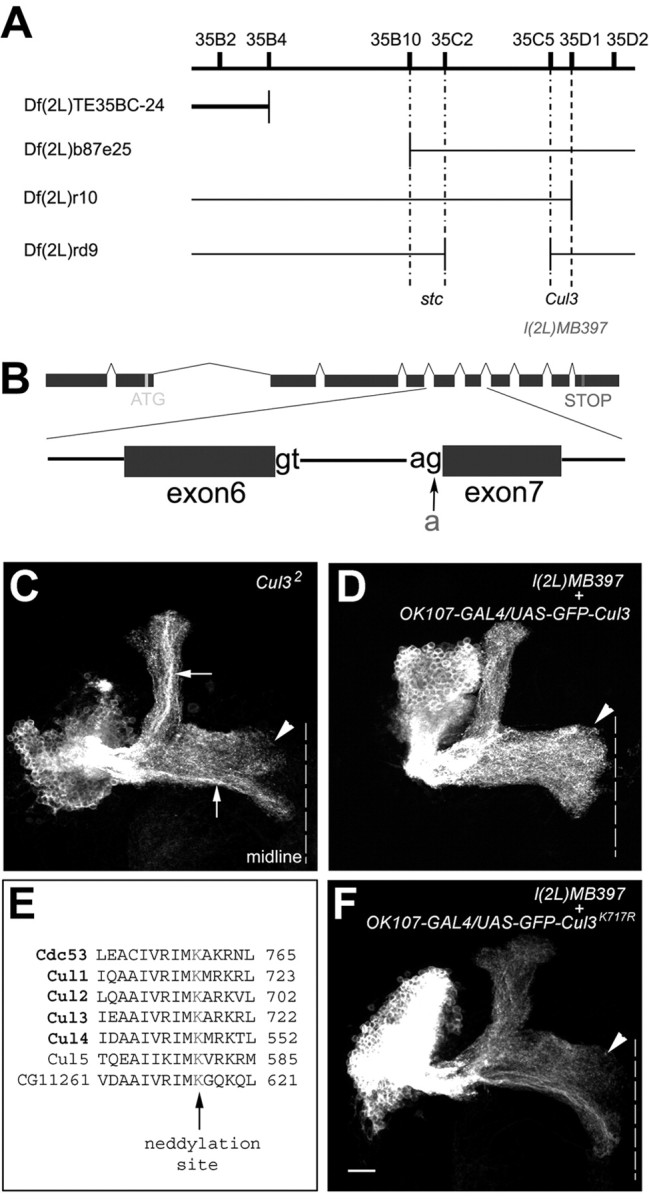
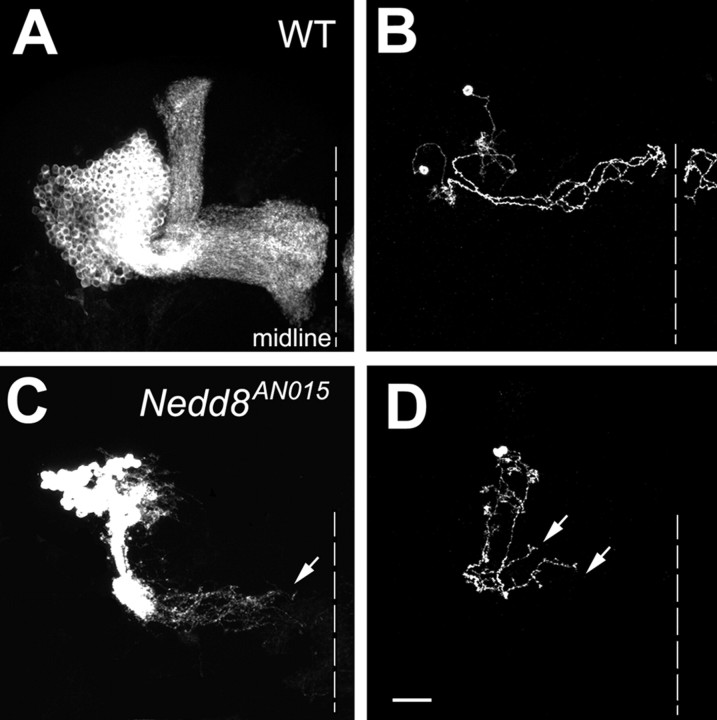
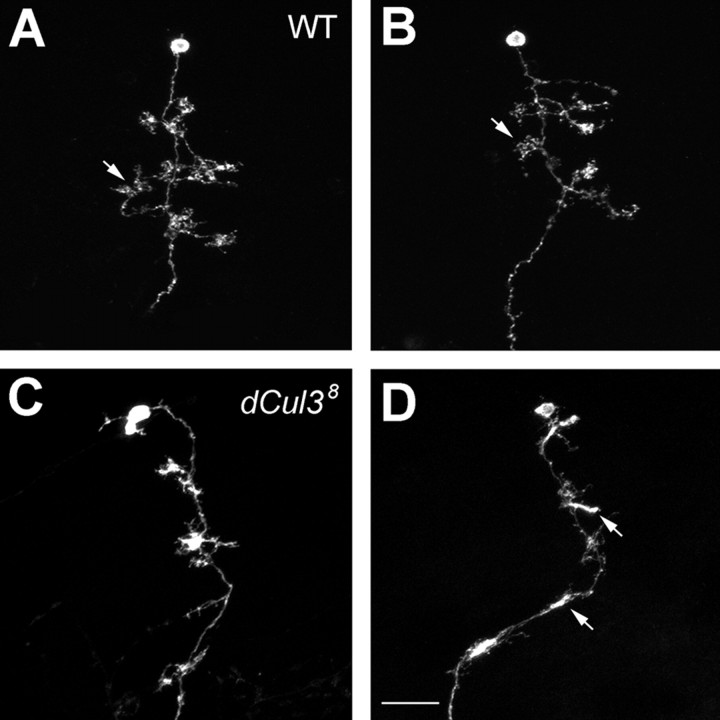
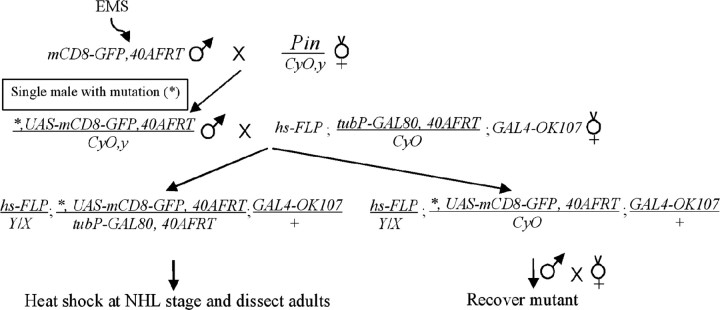
Cul3 belongs to the family of cullin proteins, which function as scaffold proteins of E3 ubiquitin ligase complexes. Here we show cell-autonomous involvement of Cul3 in axonal arborization and dendritic elaboration of Drosophila mushroom body neurons. Cul3 mutant neurons are defective in terminal morphogenesis of neurites. Interestingly, mutant axons often terminate around branching points. In addition, dendritic elaboration is severely affected in Cul3 mutant neurons. However, loss of Cul3 function does not affect extension of the axons that rarely arborize. Function of cullin-type proteins has been shown to require covalent attachment of Nedd8 (neural precursor cell-expressed developmentally downregulated), a ubiquitin-like protein. Consistent with this notion, Cul3 is inactivated by a mutation in its conserved neddylation site, and Nedd8 mutant neurons exhibit similar neuronal morphogenetic defects. Together, Cul3 plays an essential role in both axonal arborization and proper elaboration of dendrites and may require neddylation for its proper function.
Figures
Similar articles
-
The human Dcn1-like protein DCNL3 promotes Cul3 neddylation at membranes.Proc Natl Acad Sci U S A. 2009 Jul 28;106(30):12365-70. doi: 10.1073/pnas.0812528106. Epub 2009 Jul 14. Proc Natl Acad Sci U S A. 2009. PMID: 19617556 Free PMC article.
-
Drosophila Cand1 regulates Cullin3-dependent E3 ligases by affecting the neddylation of Cullin3 and by controlling the stability of Cullin3 and adaptor protein.Dev Biol. 2010 Oct 15;346(2):247-57. doi: 10.1016/j.ydbio.2010.07.031. Epub 2010 Aug 4. Dev Biol. 2010. PMID: 20691177
-
The dachshund gene is required for the proper guidance and branching of mushroom body axons in Drosophila melanogaster.J Neurobiol. 2005 Aug;64(2):133-44. doi: 10.1002/neu.20130. J Neurobiol. 2005. PMID: 15818552
-
Branching out: mechanisms of dendritic arborization.Nat Rev Neurosci. 2010 May;11(5):316-28. doi: 10.1038/nrn2836. Nat Rev Neurosci. 2010. PMID: 20404840 Free PMC article. Review.
-
Axon and dendrite pruning in Drosophila.Curr Opin Neurobiol. 2014 Aug;27:192-8. doi: 10.1016/j.conb.2014.04.005. Epub 2014 May 4. Curr Opin Neurobiol. 2014. PMID: 24793180 Free PMC article. Review.
Cited by
-
The Cullin3 ubiquitin ligase functions as a Nedd8-bound heterodimer.Mol Biol Cell. 2007 Mar;18(3):899-909. doi: 10.1091/mbc.e06-06-0542. Epub 2006 Dec 27. Mol Biol Cell. 2007. PMID: 17192413 Free PMC article.
-
Autism risk gene Cul3 alters neuronal morphology via caspase-3 activity in mouse hippocampal neurons.Front Cell Neurosci. 2024 May 9;18:1320784. doi: 10.3389/fncel.2024.1320784. eCollection 2024. Front Cell Neurosci. 2024. PMID: 38803442 Free PMC article.
-
Neuronal knockdown of Cullin3 as a Drosophila model of autism spectrum disorder.Sci Rep. 2024 Jan 17;14(1):1541. doi: 10.1038/s41598-024-51657-9. Sci Rep. 2024. PMID: 38233464 Free PMC article.
-
A missense mutation in KCTD17 causes autosomal dominant myoclonus-dystonia.Am J Hum Genet. 2015 Jun 4;96(6):938-47. doi: 10.1016/j.ajhg.2015.04.008. Epub 2015 May 14. Am J Hum Genet. 2015. PMID: 25983243 Free PMC article.
-
Identification of Genes that Maintain Behavioral and Structural Plasticity during Sleep Loss.Front Neural Circuits. 2017 Oct 23;11:79. doi: 10.3389/fncir.2017.00079. eCollection 2017. Front Neural Circuits. 2017. PMID: 29109678 Free PMC article.
References
-
- Acebes A, Ferrus A (2000) Cellular and molecular features of axon collaterals and dendrites. Trends Neurosci 23: 557-565. - PubMed
-
- Amir RE, Iwai K, Ciechanover A (2002) The NEDD8 pathway is essential for SCF(beta-TrCP)-mediated ubiquitination and processing of the NF-kappa B precursor p105. J Biol Chem 277: 23253-23259. - PubMed
-
- Arcaro KF, Lnenicka GA (1995) Intrinsic differences in axonal growth from crayfish fast and slow motoneurons. Dev Biol 168: 272-283. - PubMed
-
- Caroni P (1997) Intrinsic neuronal determinants that promote axonal sprouting and elongation. BioEssays 19: 767-775. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous