

Induction of Mycobacterium tuberculosis-specific primary and secondary T-cell responses in interleukin-15-deficient mice
- PMID: 15845497
- PMCID: PMC1087383
- DOI: 10.1128/IAI.73.5.2910-2922.2005
Induction of Mycobacterium tuberculosis-specific primary and secondary T-cell responses in interleukin-15-deficient mice
Abstract
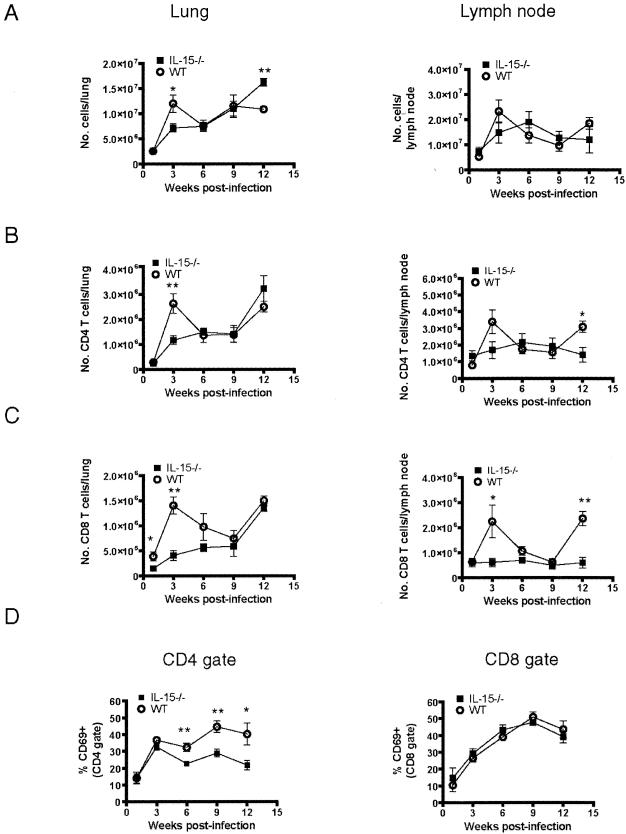
Several studies have provided evidence that interleukin-15 (IL-15) can enhance protective immune responses against Mycobacterium tuberculosis infection. However, the effects of IL-15 deficiency on the functionality of M. tuberculosis-specific CD4 and CD8 T cells are unknown. In this study, we investigated the generation and maintenance of effector and memory T-cell responses following M. tuberculosis infection of IL-15(-/-) mice. IL-15(-/-) mice had slightly higher bacterial numbers during chronic infection, which were accompanied by an increase in gamma interferon (IFN-gamma)-producing CD4 and CD8 T cells. There was no evidence of increased apoptosis or a defect in proliferation of CD8 effector T cells following M. tuberculosis infection. The induction of cytotoxic and IFN-gamma CD8 T-cell responses was normal in the absence of IL-15 signaling. The infiltration of CD4 and CD8 T cells into the lungs of "immune" IL-15(-/-) mice was delayed in response to M. tuberculosis challenge. These findings demonstrate that efficient effector CD4 and CD8 T cells can be developed following M. tuberculosis infection in the absence of IL-15 but that recall T-cell responses may be impaired.
Figures






Similar articles
-
Mycobacterium tuberculosis PstS1 amplifies IFN-γ and induces IL-17/IL-22 responses by unrelated memory CD4+ T cells via dendritic cell activation.Eur J Immunol. 2013 Sep;43(9):2386-97. doi: 10.1002/eji.201243245. Epub 2013 Jul 15. Eur J Immunol. 2013. PMID: 23719937
-
Evaluation of profile and functionality of memory T cells in pulmonary tuberculosis.Immunol Lett. 2017 Dec;192:52-60. doi: 10.1016/j.imlet.2017.10.014. Epub 2017 Oct 26. Immunol Lett. 2017. PMID: 29106984
-
Gamma interferon responses of CD4 and CD8 T-cell subsets are quantitatively different and independent of each other during pulmonary Mycobacterium bovis BCG infection.Infect Immun. 2007 May;75(5):2244-52. doi: 10.1128/IAI.00024-07. Epub 2007 Feb 16. Infect Immun. 2007. PMID: 17307945 Free PMC article.
-
Functions of T-cell subsets and cytokines in mycobacterial infections.Eur Respir J Suppl. 1995 Sep;20:668s-675s. Eur Respir J Suppl. 1995. PMID: 8590567 Review.
-
Type I, II, and III Interferons: Regulating Immunity to Mycobacterium tuberculosis Infection.Arch Immunol Ther Exp (Warsz). 2016 Feb;64(1):19-31. doi: 10.1007/s00005-015-0365-7. Epub 2015 Sep 11. Arch Immunol Ther Exp (Warsz). 2016. PMID: 26362801 Review.
Cited by
-
Limited recognition of Mycobacterium tuberculosis-infected macrophages by polyclonal CD4 and CD8 T cells from the lungs of infected mice.Mucosal Immunol. 2020 Jan;13(1):140-148. doi: 10.1038/s41385-019-0217-6. Epub 2019 Oct 21. Mucosal Immunol. 2020. PMID: 31636345 Free PMC article.
-
Impact of toll-like receptor 2 deficiency on immune responses to mycobacterial antigens.Infect Immun. 2011 Nov;79(11):4649-56. doi: 10.1128/IAI.05724-11. Epub 2011 Aug 15. Infect Immun. 2011. PMID: 21844233 Free PMC article.
-
Aberrant TGF-beta signaling reduces T regulatory cells in ICAM-1-deficient mice, increasing the inflammatory response to Mycobacterium tuberculosis.J Leukoc Biol. 2009 Sep;86(3):713-25. doi: 10.1189/jlb.1208740. Epub 2009 May 19. J Leukoc Biol. 2009. PMID: 19454651 Free PMC article.
-
Development of a secondary immune response to Mycobacterium tuberculosis is independent of Toll-like receptor 2.Infect Immun. 2011 Mar;79(3):1118-23. doi: 10.1128/IAI.01076-10. Epub 2010 Dec 20. Infect Immun. 2011. PMID: 21173309 Free PMC article.
-
Trans-presentation of interleukin-15 by interleukin-15 receptor alpha is dispensable for the pathogenesis of autoimmune type 1 diabetes.Cell Mol Immunol. 2017 Jul;14(7):590-596. doi: 10.1038/cmi.2015.102. Epub 2016 Feb 8. Cell Mol Immunol. 2017. PMID: 26853723 Free PMC article.
References
-
- Avice, M. N., C. E. Demeure, G. Delespesse, M. Rubio, M. Armant, and M. Sarfati. 1998. IL-15 promotes IL-12 production by human monocytes via T cell-dependent contact and may contribute to IL-12-mediated IFN-gamma secretion by CD4+ T cells in the absence of TCR ligation. J. Immunol. 161:3408-3415. - PubMed
-
- Berard, M., K. Brandt, S. Bulfone-Paus, and D. F. Tough. 2003. IL-15 promotes the survival of naive and memory phenotype CD8+ T cells. J. Immunol. 170:5018-5026. - PubMed
-
- Doherty, T. M., R. A. Seder, and A. Sher. 1996. Induction and regulation of IL-15 expression in murine macrophages. J. Immunol. 156:735-741. - PubMed
-
- Giulietti, A., L. Overbergh, D. Valckx, B. Decallonne, R. Bouillon, and C. Mathieu. 2001. An overview of real-time quantitative PCR: applications to quantify cytokine gene expression. Methods 25:386-401. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
