

Disulfide bridge formation between SecY and a translocating polypeptide localizes the translocation pore to the center of SecY
- PMID: 15851514
- PMCID: PMC2171872
- DOI: 10.1083/jcb.200412019
Disulfide bridge formation between SecY and a translocating polypeptide localizes the translocation pore to the center of SecY
Abstract
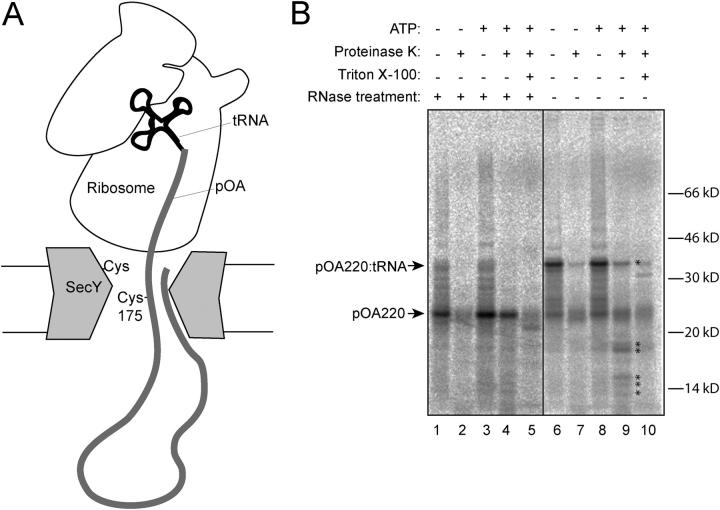
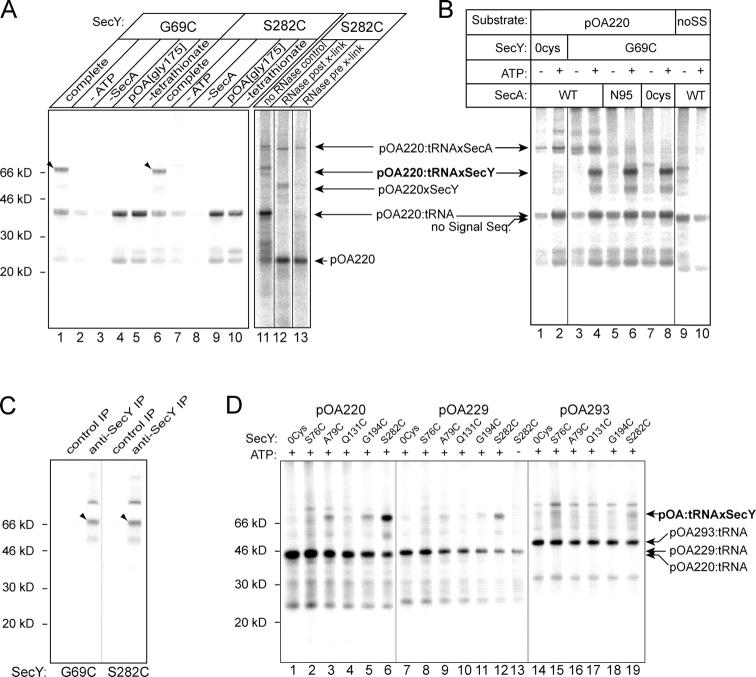
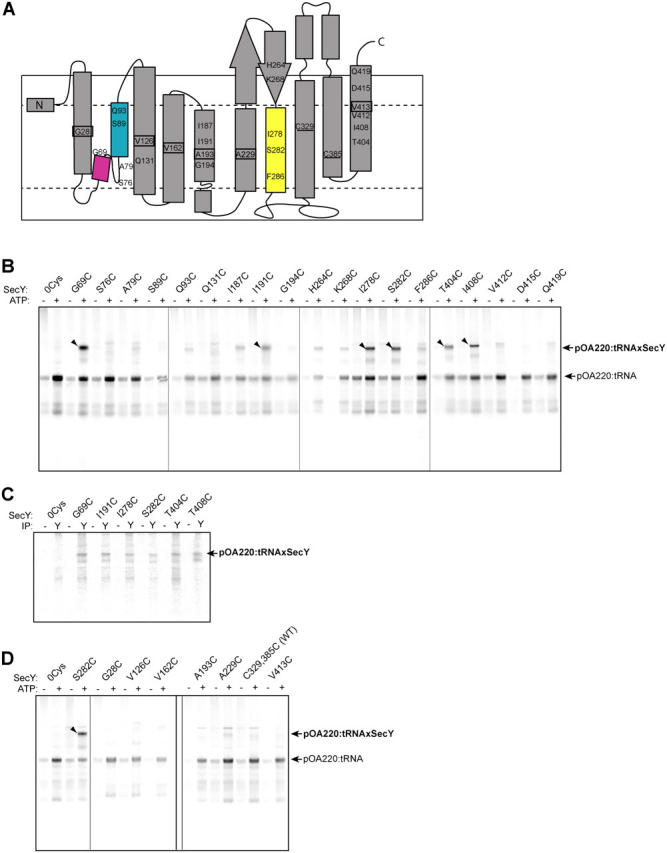
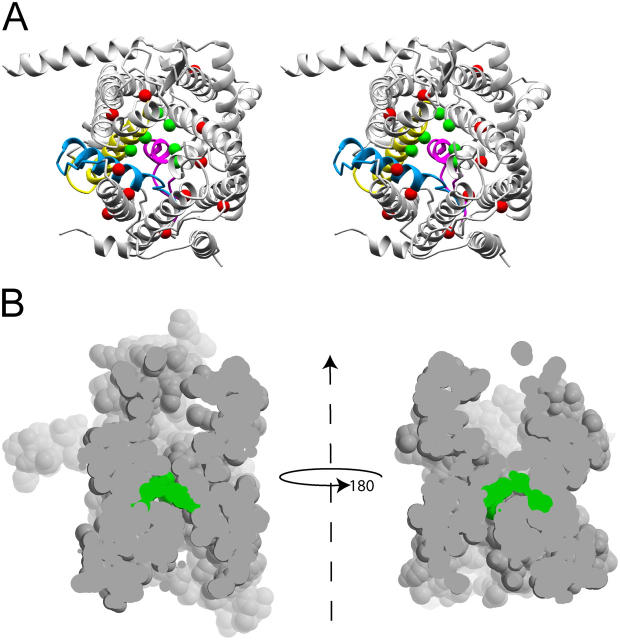
During their biosynthesis, many proteins pass through the membrane via a hydrophilic channel formed by the heterotrimeric Sec61/SecY complex. Whether this channel forms at the interface of multiple copies of Sec61/SecY or is intrinsic to a monomeric complex, as suggested by the recently solved X-ray structure of the Methanococcus jannaschii SecY complex, is a matter of contention. By introducing a single cysteine at various positions in Escherichia coli SecY and testing its ability to form a disulfide bond with a single cysteine in a translocating chain, we provide evidence that translocating polypeptides pass through the center of the SecY complex. The strongest cross-links were observed with residues that would form a constriction in an hourglass-shaped pore. This suggests that the channel makes only limited contact with a translocating polypeptide, thus minimizing the energy required for translocation.
Figures
References
-
- Bankaitis, V., B. Rasmussen, and P. Bassford. 1984. Intragenic suppressor mutations that restore export of maltose-binding protein with a truncated signal peptide. Cell. 37:243–252. - PubMed
-
- Beckmann, R., D. Bubeck, R. Grassucci, P. Penczek, A. Verschoor, G. Blobel, and J. Frank. 1997. Alignment of conduits for the nascent polypeptide chain in the ribosome-Sec61 complex. Science. 278:2123–2126. - PubMed
-
- Crowley, K.S., S.R. Liao, V.E. Worrell, G.D. Reinhart, and A.E. Johnson. 1994. Secretory proteins move through the endoplasmic reticulum membrane via an aqueous, gated pore. Cell. 78:461–471. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
