

Evolutionary origin of cAMP-based chemoattraction in the social amoebae
- PMID: 15851658
- PMCID: PMC1088387
- DOI: 10.1073/pnas.0502238102
Evolutionary origin of cAMP-based chemoattraction in the social amoebae
Abstract
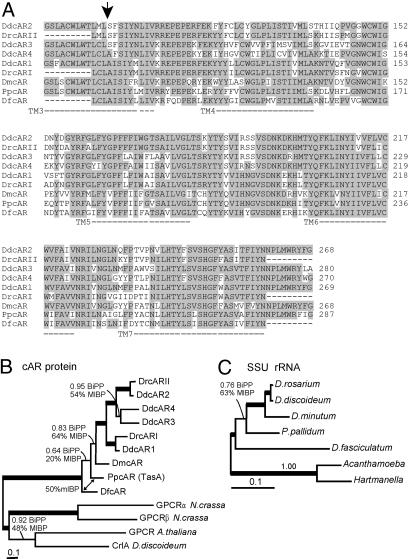
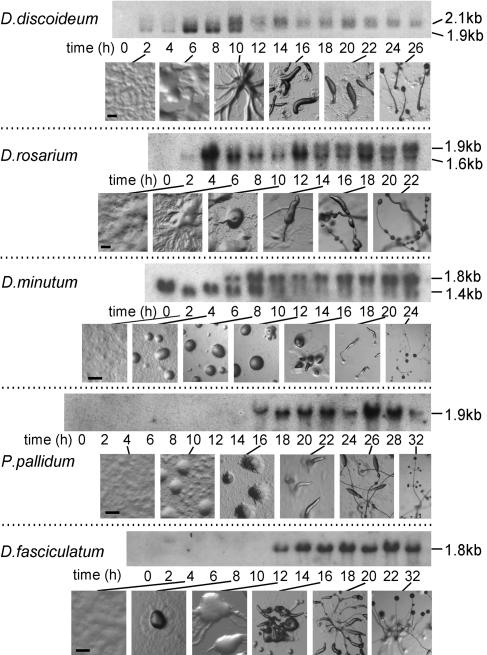
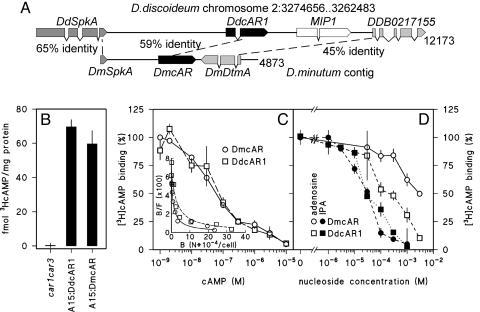
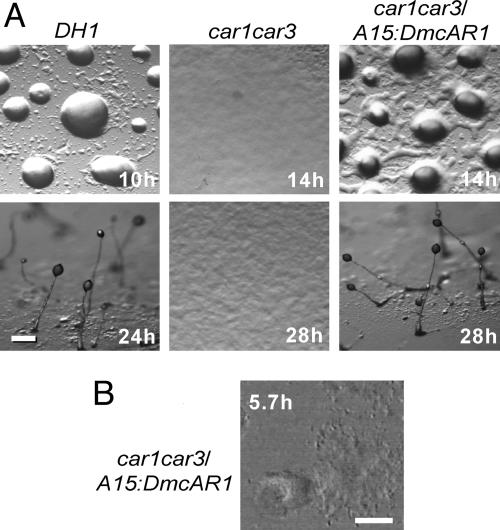
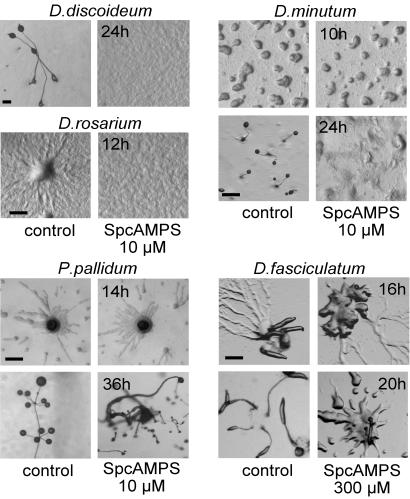
Phenotypic novelties can arise if integrated developmental pathways are expressed at new developmental stages and then recruited to serve new functions. We analyze the origin of a novel developmental trait of Dictyostelid amoebae: the evolution of cAMP as a developmental chemoattractant. We show that cAMP's role of attracting starving amoebae arose through recruitment of a pathway that originally evolved to coordinate fruiting body morphogenesis. Orthologues of the high-affinity cAMP receptor (cAR), cAR1, were identified in a selection of species that span the Dictyostelid phylogeny. The cAR1 orthologue from the basal species Dictyostelium minutum restored aggregation and development when expressed in an aggregation-defective mutant of the derived species Dictyostelium discoideum that lacks high-affinity cARs, thus demonstrating that the D. minutum cAR is a fully functional cAR. cAR1 orthologues from basal species are expressed during fruiting body formation, and only this process, and not aggregation, was disrupted by abrogation of cAR1 function. This is in contrast to derived species, where cAR1 is also expressed during aggregation and critically regulates this process. Our data show that coordination of fruiting body formation is the ancestral function of extracellular cAMP signaling, whereas its derived role in aggregation evolved by recruitment of a preexisting pathway to an earlier stage of development. This most likely occurred by addition of distal cis-regulatory regions to existing cAMP signaling genes.
Figures
References
-
- Long, M., Betran, E., Thornton, K. & Wang, W. (2003) Nat. Rev. Genet. 4, 865-875. - PubMed
-
- True, J. R. & Carroll, S. B. (2002) Annu. Rev. Cell Dev. Biol. 18, 53-80. - PubMed
-
- Lee, P. N., Callaerts, P., de Couet, H. G. & Martindale, M. Q. (2003) Nature 424, 1061-1065. - PubMed
-
- Irish, V. F. (2003) BioEssays 25, 637-646. - PubMed
-
- Raper, K. B. (1984) The Dictyostelids (Princeton Univ. Press, Princeton).
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
