

Evaluating putative mechanisms of the mitotic spindle checkpoint
- PMID: 15851663
- PMCID: PMC1088361
- DOI: 10.1073/pnas.0409142102
Evaluating putative mechanisms of the mitotic spindle checkpoint
Abstract
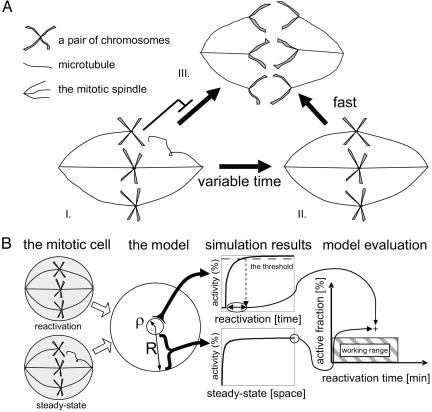
The mitotic spindle checkpoint halts the cell cycle until all chromosomes are attached to the mitotic spindles. Evidence suggests that the checkpoint prevents cell-cycle progression by inhibiting the activity of the APC-Cdc20 complex, but the precise mechanism underlying this inhibition is not yet known. Here, we use mathematical modeling to compare several mechanisms that could account for this inhibition. We describe the interplay between the capacities to strongly inhibit cell-cycle progression before spindle attachment on one hand and to rapidly resume cell-cycle progression once the last kinetochore is attached on the other hand. We find that inhibition that is restricted to the kinetochore region is not sufficient for supporting both requirements when realistic diffusion constants are considered. A mechanism that amplifies the checkpoint signal through autocatalyzed inhibition is also insufficient. In contrast, amplifying the signal through the release of a diffusible inhibitory complex can support reliable checkpoint function. Our results suggest that the design of the spindle checkpoint network is limited by physical constraints imposed by realistic diffusion constants and the relevant spatial and temporal dimensions where computation is performed.
Figures


References
-
- Hartwell, L. H. & Weinert, T. A. (1989) Science 246, 629-634. - PubMed
-
- Kastan, M. B. & Bartek, J. (2004) Nature 432, 316-323. - PubMed
-
- Hernando, E., Nahle, Z., Juan, G., Diaz-Rodriguez, E., Alaminos, M., Hemann, M., Michel, L., Mittal, V., Gerald, W., Benezra, R., et al. (2004) Nature 430, 797-802. - PubMed
-
- Rajagopalan, H. & Lengauer, C. (2004) Nature 432, 338-341. - PubMed
-
- Shonn, M. A., McCarroll, R. & Murray, A. W. (2000) Science 289, 300-303. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
