

Role of CXCR2/CXCR2 ligands in vascular remodeling during bronchiolitis obliterans syndrome
- PMID: 15864347
- PMCID: PMC1087179
- DOI: 10.1172/JCI24233
Role of CXCR2/CXCR2 ligands in vascular remodeling during bronchiolitis obliterans syndrome
Abstract
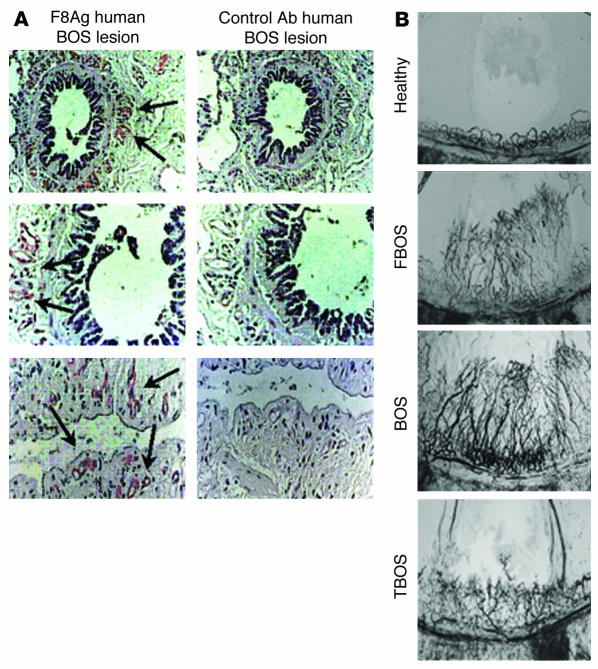
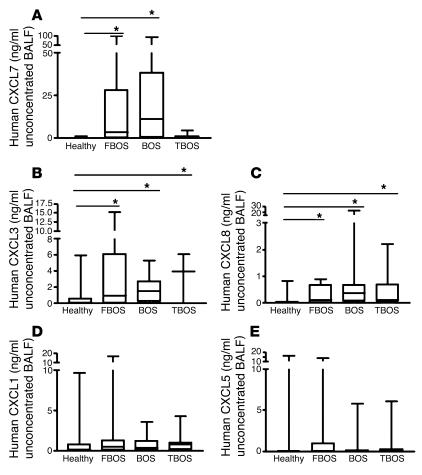
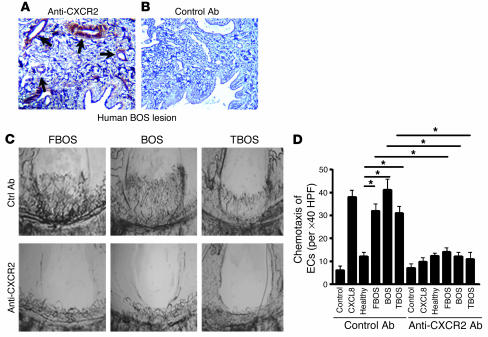
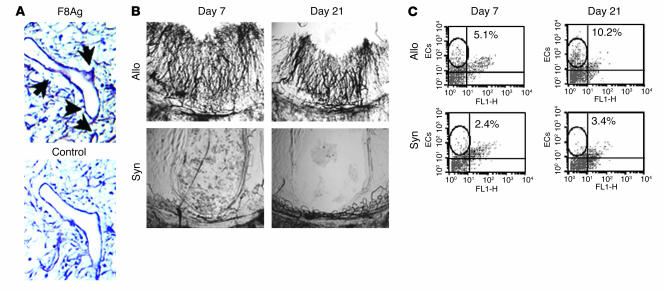
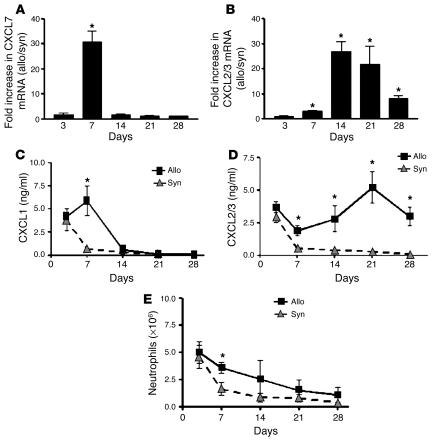
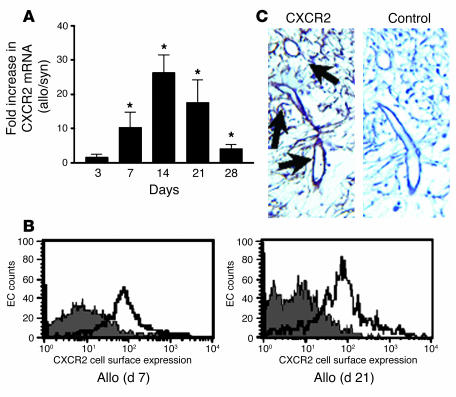
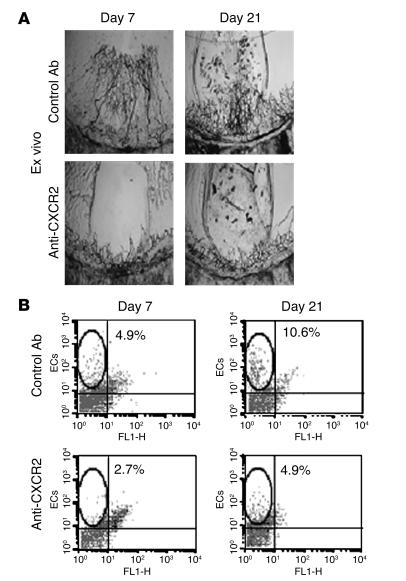
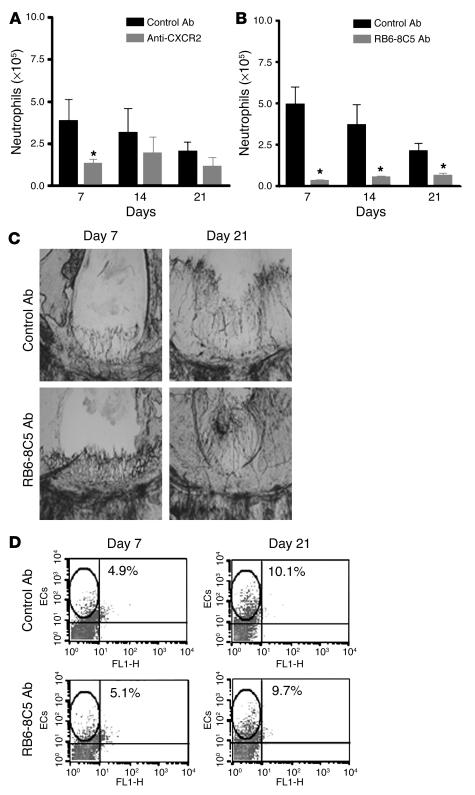
Angiogenesis and vascular remodeling support fibroproliferative processes; however, no study has addressed the importance of angiogenesis during fibro-obliteration of the allograft airway during bronchiolitis obliterans syndrome (BOS) that occurs after lung transplantation. The ELR(+) CXC chemokines both mediate neutrophil recruitment and promote angiogenesis. Their shared endothelial cell receptor is the G-coupled protein receptor CXC chemokine receptor 2 (CXCR2). We found that elevated levels of multiple ELR(+) CXC chemokines correlated with the presence of BOS. Proof-of-concept studies using a murine model of BOS not only demonstrated an early neutrophil infiltration but also marked vascular remodeling in the tracheal allografts. In addition, tracheal allograft ELR(+) CXC chemokines were persistently expressed even in the absence of significant neutrophil infiltration and were temporally associated with vascular remodeling during fibro-obliteration of the tracheal allograft. Furthermore, in neutralizing studies, treatment with anti-CXCR2 Abs inhibited early neutrophil infiltration and later vascular remodeling, which resulted in the attenuation of murine BOS. A more profound attenuation of fibro-obliteration was seen when CXCR2(-/-) mice received cyclosporin A. This supports the notion that the CXCR2/CXCR2 ligand biological axis has a bimodal function during the course of BOS: early, it is important for neutrophil recruitment and later, during fibro-obliteration, it is important for vascular remodeling independent of neutrophil recruitment.
Figures
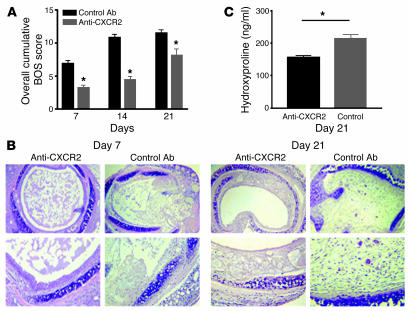
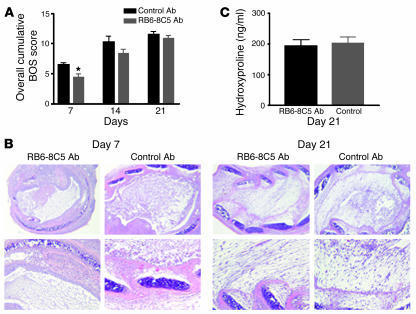

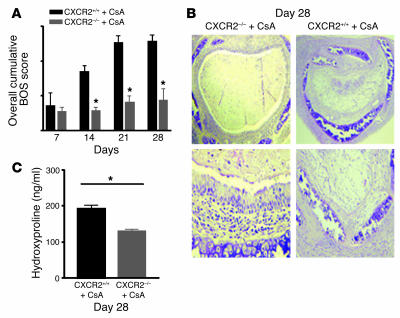
Comment in
-
Chemokine-mediated angiogenesis: an essential link in the evolution of airway fibrosis?J Clin Invest. 2005 May;115(5):1133-6. doi: 10.1172/JCI25193. J Clin Invest. 2005. PMID: 15864341 Free PMC article.
References
-
- Trulock EP. Lung transplantation. Am. J. Respir. Crit. Care Med. 1997;155:789–818. - PubMed
-
- Arcasoy SM, Kotloff RM. Lung transplantation. N. Engl. J. Med. 1999;340:1081–1091. - PubMed
-
- Folkman J. Angiogenesis in cancer, vascular, rheumatoid and other disease. Nat. Med. 1995;1:27–31. - PubMed
-
- Belperio JA, et al. CXC chemokines in angiogenesis. J. Leukoc. Biol. 2000;68:1–8. - PubMed
-
- Strieter RM, Belperio JA, Keane MP. CXC chemokines in vascular remodeling related to pulmonary fibrosis. Am. J. Respir. Cell Mol. Biol. 2003;29(Suppl. 3):S67–S69. - PubMed
