

Biochemical properties of purified human retinol dehydrogenase 12 (RDH12): catalytic efficiency toward retinoids and C9 aldehydes and effects of cellular retinol-binding protein type I (CRBPI) and cellular retinaldehyde-binding protein (CRALBP) on the oxidation and reduction of retinoids
- PMID: 15865448
- PMCID: PMC2679700
- DOI: 10.1021/bi050226k
Biochemical properties of purified human retinol dehydrogenase 12 (RDH12): catalytic efficiency toward retinoids and C9 aldehydes and effects of cellular retinol-binding protein type I (CRBPI) and cellular retinaldehyde-binding protein (CRALBP) on the oxidation and reduction of retinoids
Abstract
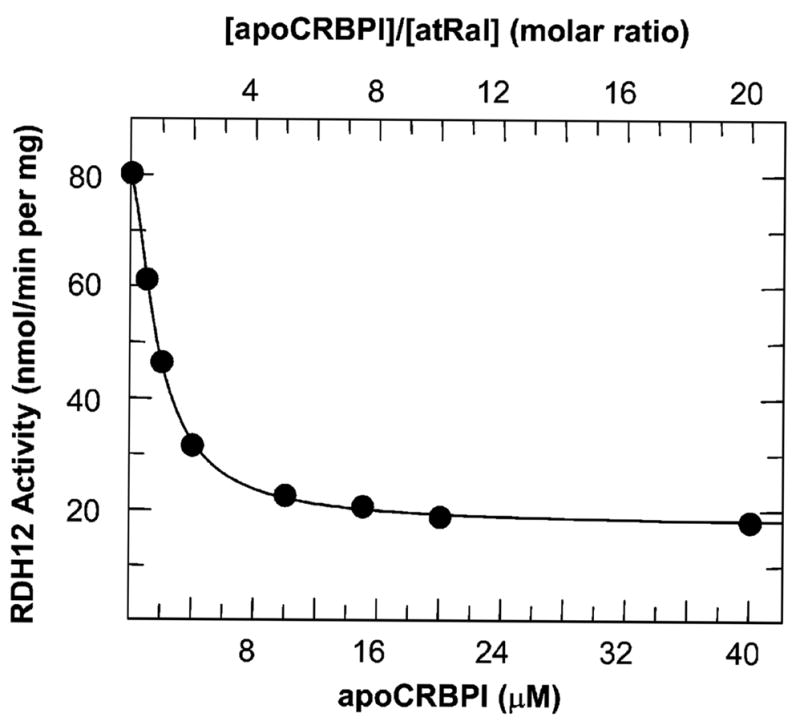
Retinol dehydrogenase 12 (RDH12) is a novel member of the short-chain dehydrogenase/reductase superfamily of proteins that was recently linked to Leber's congenital amaurosis 3 (LCA). We report the first biochemical characterization of purified human RDH12 and analysis of its expression in human tissues. RDH12 exhibits approximately 2000-fold lower K(m) values for NADP(+) and NADPH than for NAD(+) and NADH and recognizes both retinoids and lipid peroxidation products (C(9) aldehydes) as substrates. The k(cat) values of RDH12 for retinaldehydes and C(9) aldehydes are similar, but the K(m) values are, in general, lower for retinoids. The enzyme exhibits the highest catalytic efficiency for all-trans-retinal (k(cat)/K(m) approximately 900 min(-)(1) microM(-)(1)), followed by 11-cis-retinal (450 min(-)(1) mM(-)(1)) and 9-cis-retinal (100 min(-)(1) mM(-)(1)). Analysis of RDH12 activity toward retinoids in the presence of cellular retinol-binding protein (CRBP) type I or cellular retinaldehyde-binding protein (CRALBP) suggests that RDH12 utilizes the unbound forms of all-trans- and 11-cis-retinoids. As a result, the widely expressed CRBPI, which binds all-trans-retinol with much higher affinity than all-trans-retinaldehyde, restricts the oxidation of all-trans-retinol by RDH12, but has little effect on the reduction of all-trans-retinaldehyde, and CRALBP inhibits the reduction of 11-cis-retinal stronger than the oxidation of 11-cis-retinol, in accord with its higher affinity for 11-cis-retinal. Together, the tissue distribution of RDH12 and its catalytic properties suggest that, in most tissues, RDH12 primarily contributes to the reduction of all-trans-retinaldehyde; however, at saturating concentrations of peroxidic aldehydes in the cells undergoing oxidative stress, for example, photoreceptors, RDH12 might also play a role in detoxification of lipid peroxidation products.
Figures






Similar articles
-
Effect of lipid peroxidation products on the activity of human retinol dehydrogenase 12 (RDH12) and retinoid metabolism.Biochim Biophys Acta. 2008 Jun;1782(6):421-5. doi: 10.1016/j.bbadis.2008.03.004. Epub 2008 Mar 18. Biochim Biophys Acta. 2008. PMID: 18396173 Free PMC article.
-
Overproduction of bioactive retinoic acid in cells expressing disease-associated mutants of retinol dehydrogenase 12.J Biol Chem. 2007 Dec 7;282(49):35621-8. doi: 10.1074/jbc.M706372200. Epub 2007 Oct 9. J Biol Chem. 2007. PMID: 17925390
-
Retinoid processing proteins in the ocular ciliary epithelium.Mol Vis. 2005 May 18;11:356-65. Mol Vis. 2005. PMID: 15928609
-
Medium- and short-chain dehydrogenase/reductase gene and protein families : Medium-chain and short-chain dehydrogenases/reductases in retinoid metabolism.Cell Mol Life Sci. 2008 Dec;65(24):3936-49. doi: 10.1007/s00018-008-8591-3. Cell Mol Life Sci. 2008. PMID: 19011747 Free PMC article. Review.
-
Enzymes and binding proteins affecting retinoic acid concentrations.J Steroid Biochem Mol Biol. 1995 Jun;53(1-6):497-502. doi: 10.1016/0960-0760(95)00096-i. J Steroid Biochem Mol Biol. 1995. PMID: 7626500 Review.
Cited by
-
Novel Heterozygous Deletion in Retinol Dehydrogenase 12 (RDH12) Causes Familial Autosomal Dominant Retinitis Pigmentosa.Front Genet. 2020 Apr 8;11:335. doi: 10.3389/fgene.2020.00335. eCollection 2020. Front Genet. 2020. PMID: 32322264 Free PMC article.
-
Dynamic lipid turnover in photoreceptors and retinal pigment epithelium throughout life.Prog Retin Eye Res. 2022 Jul;89:101037. doi: 10.1016/j.preteyeres.2021.101037. Epub 2021 Dec 29. Prog Retin Eye Res. 2022. PMID: 34971765 Free PMC article. Review.
-
Characterization of key residues and membrane association domains in retinol dehydrogenase 10.Biochem J. 2009 Apr 1;419(1):113-22, 1 p following 122. doi: 10.1042/BJ20080812. Biochem J. 2009. PMID: 19102727 Free PMC article.
-
Functions of Intracellular Retinoid Binding-Proteins.Subcell Biochem. 2016;81:21-76. doi: 10.1007/978-94-024-0945-1_2. Subcell Biochem. 2016. PMID: 27830500 Free PMC article. Review.
-
Retinol Dehydrogenases Regulate Vitamin A Metabolism for Visual Function.Nutrients. 2016 Nov 22;8(11):746. doi: 10.3390/nu8110746. Nutrients. 2016. PMID: 27879662 Free PMC article. Review.
References
-
- Janecke AR, Thompson DA, Utermann G, Becker C, Hubner CA, Schmid E, McHenry CL, Nair AR, Ruschendorf F, Heckenlively J, Wissinger B, Nurnberg P, Gal A. Mutations in RDH12 encoding a photoreceptor cell retinol dehydrogenase cause childhood-onset severe retinal dystrophy. Nat Genet. 2004;36:850–854. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
