

Mnt-Max to Myc-Max complex switching regulates cell cycle entry
- PMID: 15866886
- PMCID: PMC2171929
- DOI: 10.1083/jcb.200411013
Mnt-Max to Myc-Max complex switching regulates cell cycle entry
Abstract
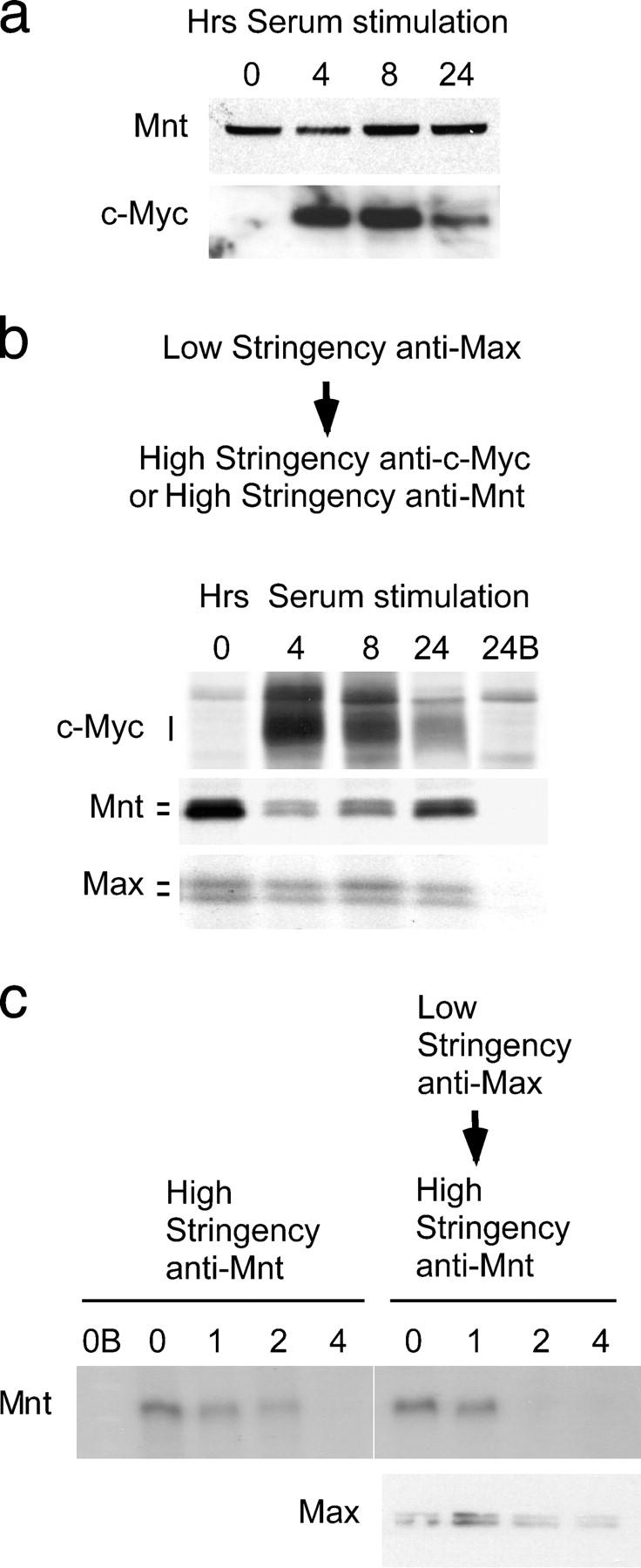
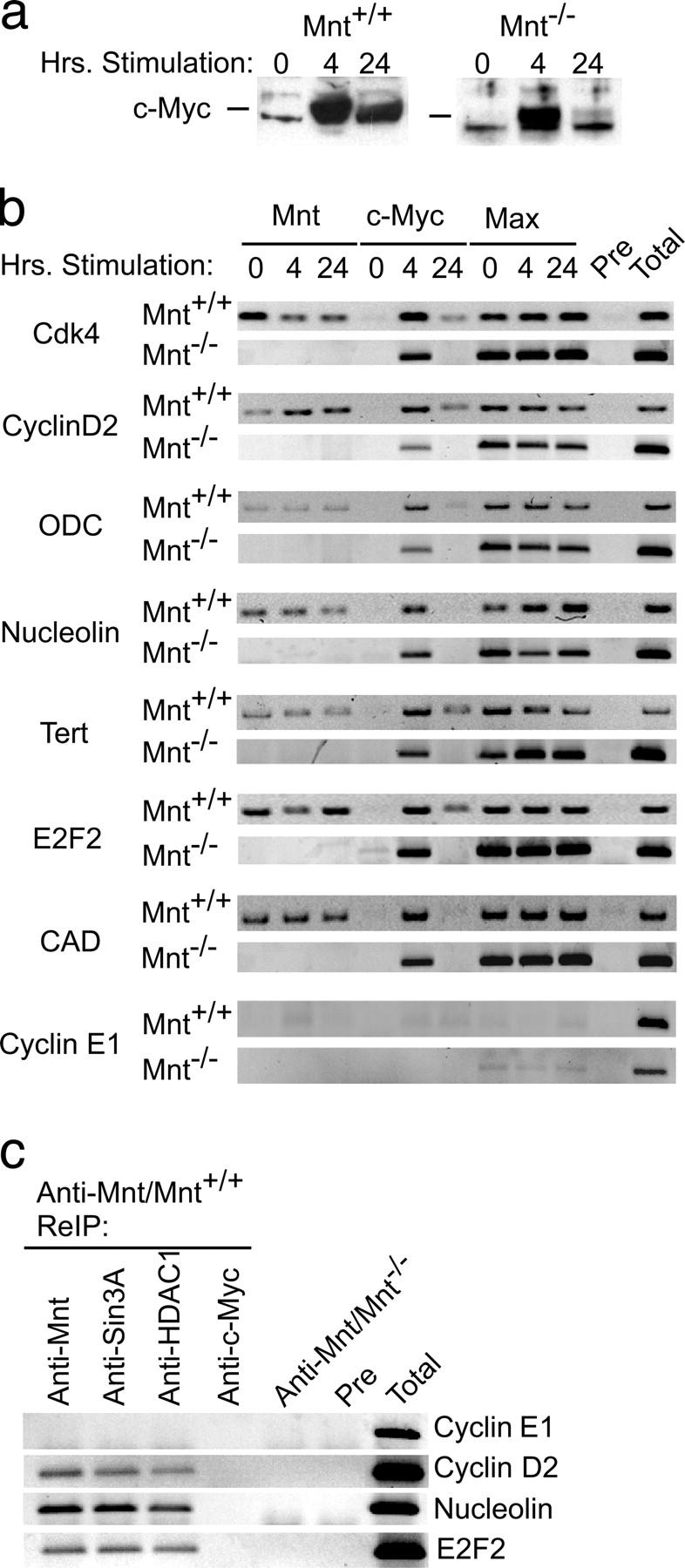
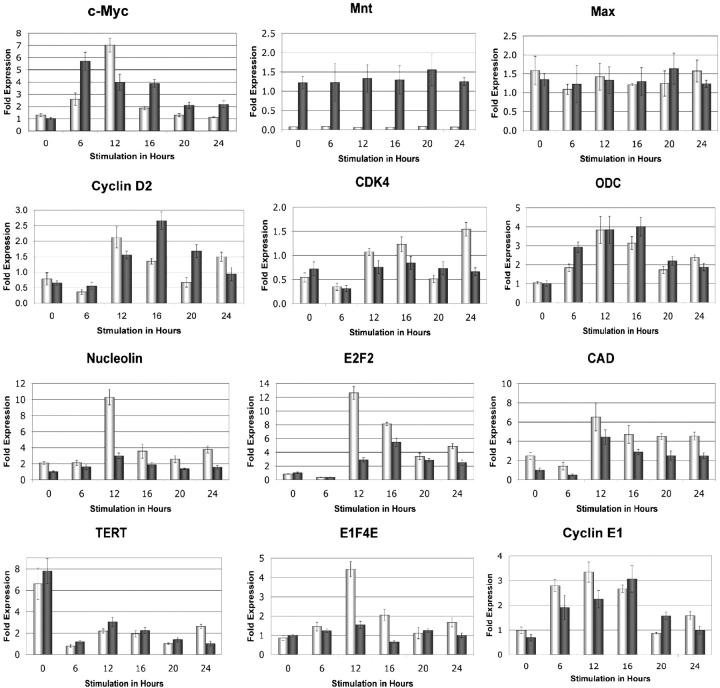
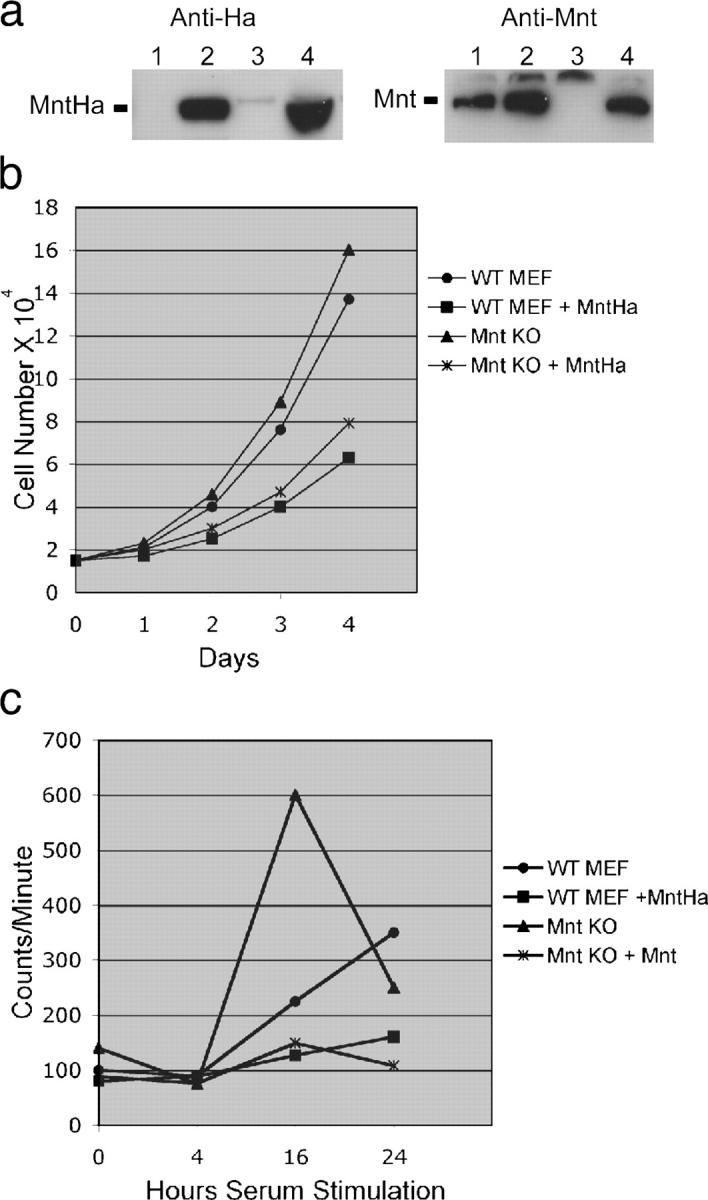
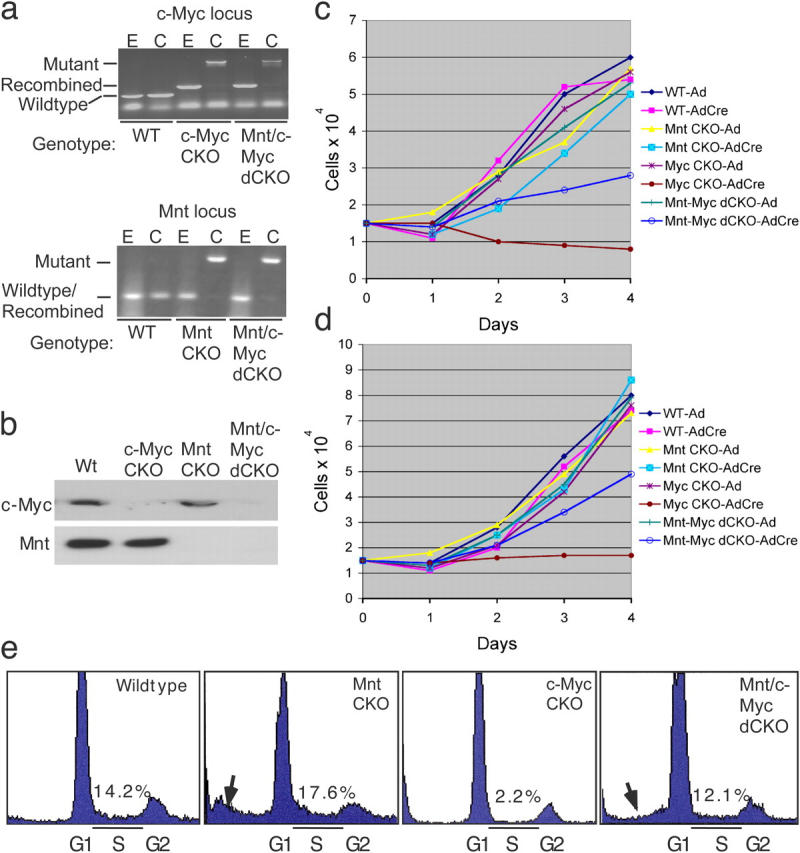
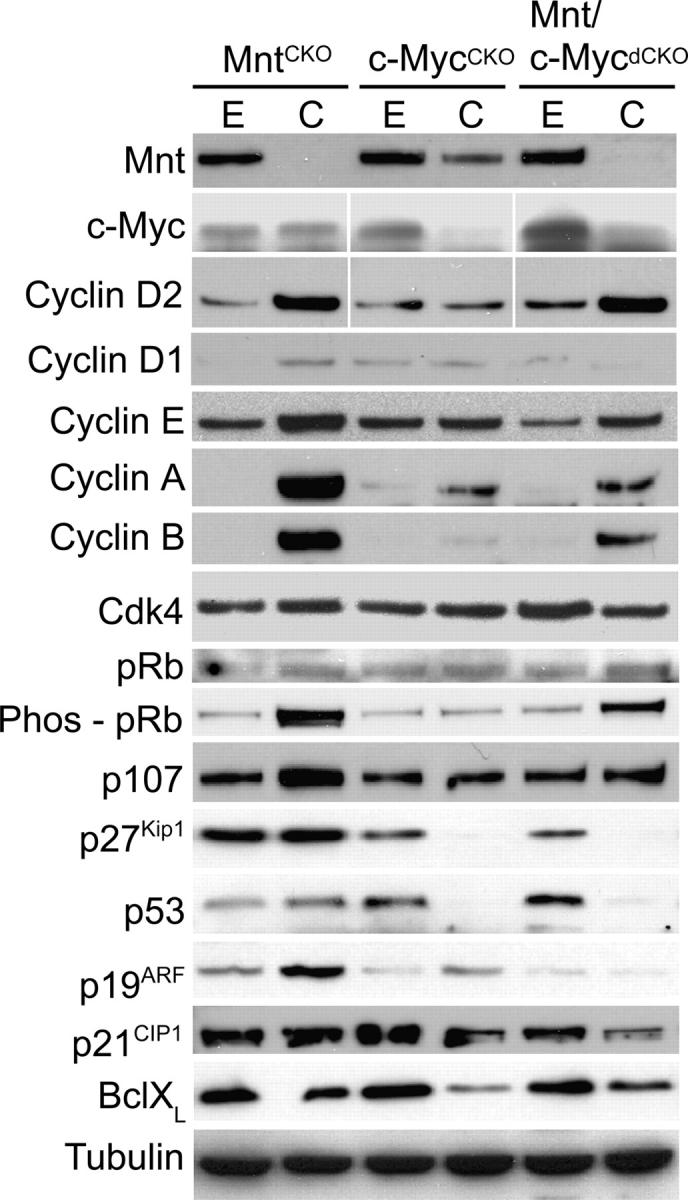
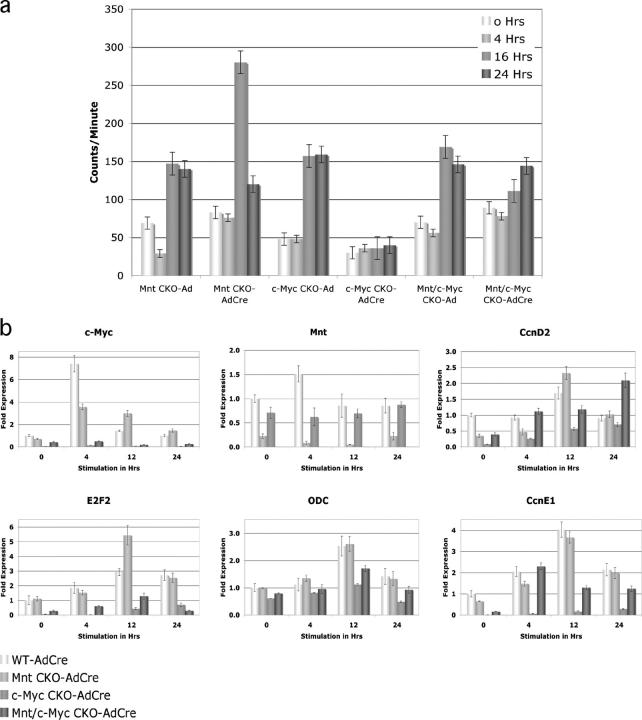
The c-Myc oncoprotein is strongly induced during the G0 to S-phase transition and is an important regulator of cell cycle entry. In contrast to c-Myc, the putative Myc antagonist Mnt is maintained at a constant level during cell cycle entry. Mnt and Myc require interaction with Max for specific DNA binding at E-box sites, but have opposing transcriptional activities. Here, we show that c-Myc induction during cell cycle entry leads to a transient decrease in Mnt-Max complexes and a transient switch in the ratio of Mnt-Max to c-Myc-Max on shared target genes. Mnt overexpression suppressed cell cycle entry and cell proliferation, suggesting that the ratio of Mnt-Max to c-Myc-Max is critical for cell cycle entry. Furthermore, simultaneous Cre-Lox mediated deletion of Mnt and c-Myc in mouse embryo fibroblasts rescued the cell cycle entry and proliferative block caused by c-Myc ablation alone. These results demonstrate that Mnt-Myc antagonism plays a fundamental role in regulating cell cycle entry and proliferation.
Figures
References
-
- Amati, B., S. Dalton, M.W. Brooks, T.D. Littlewood, G.I. Evan, and H. Land. 1992. Transcriptional activation by the human c-Myc oncoprotein in yeast requires interaction with Max. Nature. 359:423–426. - PubMed
-
- Askew, D.S., R.A. Ashmun, B.C. Simmons, and J.L. Cleveland. 1991. Constitutive c-myc expression in an IL-3-dependent myeloid cell line suppresses cell cycle arrest and accelerates apoptosis. Oncogene. 6:1915–1922. - PubMed
-
- Ayer, D.E. 1999. Histone deacetylases: transcriptional repression with SINers and NuRDs. Trends Cell Biol. 9:193–198. - PubMed
-
- Blackwood, E.M., and R.N. Eisenman. 1991. Max: a helix-loop-helix zipper protein that forms a sequence-specific DNA-binding complex with Myc. Science. 251:1211–1217. - PubMed
-
- Blackwood, E.M., B. Luscher, and R.N. Eisenman. 1992. Myc and Max associate in vivo. Genes Dev. 6:71–80. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
